|
#1
 12.09.2013, 20:32
12.09.2013, 20:32
|
||||
|
||||
|
http://www.polit.ru/article/2008/05/23/geny/
Мы публикуем полную стенограмму лекции, прочитанной кандидатом биологических наук, ведущим научным сотрудником лаборатории анализа генома Института общей генетики им. Н.И.Вавилова Светланой Александровной Боринской 17 апреля 2008 года в клубе – литературном кафе Bilingua в рамках проекта «Публичные лекции «Полит.ру»». Лекция стала продолжением цикла (начало - выступления Кирилла Еськова "Палеонтология и макроэволюция" и Михаила Гельфанда "Геномы и эволюция"), призванного познакомить с современным состоянием естествознания.  Светлана Боринская (фото Наташи Четвериковой) С тех пор, как почти 200 тысяч лет назад человек как биологический вид появился в Африке, он продолжал дело всех своих предков и успешно размножался. Человечество размножалось столь успешно, что в родной Африке места стало не хватать, и люди вышли за ее пределы, заселив Азию, Европу и, перейдя по суше, которая была вместо нынешнего Берингова пролива, в Америку, успешно расселились и там. Предполагается, что почти все ныне обитаемые уголки Земли были заняты примерно 25-15 тысяч лет назад. Таким образом, я сразу ответила на вопросы о том, где возник человек – в Африке, когда он возник, и могу сразу же сказать, как он возник: он получился из обезьяны путем естественного отбора, но то была не современная обезьяна, а общий предок человека и других ныне живущих высших приматов. Теперь все самые нетерпеливые могут расслабиться, выпить пива, потому что все основное вы уже услышали, и до конца лекции я буду излагать подробности сей истории. Прежде, чем более детально рассмотреть разные аспекты эволюции человека, я попробую осветить еще некоторые вопросы. С точки зрения генетиков, а также некоторых коллег из смежных областей науки, например, антропологии и зоологии, самым близким родственником человека является шимпанзе. Видов шимпанзе два: обыкновенный и карликовый (бонобо). Они разделились относительно недавно – 2 млн. лет назад, а до этого они примерно 6-7 млн. лет назад отделились от той ветви, который ведет к человеку. Еще раньше общие предки шимпанзе и человека отделились от предков гориллы, орангутана, а еще раньше – от предков остальных видов обезьян (слайд 1). И сегодняшняя лекция будет посвящена вопросам когда, где, как и зачем появился вид Homo sapiens. Я хочу подчеркнуть, что излагаю свое частное мнение, а не истину в последней инстанции, поэтому некоторые вопросы, которые я хочу осветить, вполне дискуссионные, и я надеюсь, что у нас останется время для их обсуждения. Для того, чтобы понять, как генетические изменения привели к появлению человека, как гены в эволюции менялись и изменения эти “сделали человека”, мы сначала рассмотрим более простой вопрос: как гены “делают человека” в индивидуальном развитии, которое мы можем непосредственно наблюдать, и изучение которого не требует проводить какие-то сложные реконструкции далекого прошлого. Итак, каким образом из зародышевой клетки получаемся мы с вами? Чтобы пояснить это, я скопировала из лекции Михаила Гельфанда слайд, который мне очень понравился, и немножко его модифицировала (слайд 2). В прошлой лекции это называлось ликбезом, это школьные знания, и я их напомню. Каждый ген имеет структурную часть, которая кодирует белок, и регуляторную часть, в которой записано, когда гену работать и когда – не работать, т.е., в каких условиях и в какие периоды развития каждой клетки этот ген следует включить, а когда – выключить. Когда ген работает, с него синтезируется молекула РНК – посредник, переносящий информацию, на рибосомы - машины, синтезирующие белки. Белки, в свою очередь, выполняют очень много работ в клетке, строят ее структурно, и выполняют разные каталитические функции. Сейчас выяснилось, что РНК выполняют тоже много функций, только у ДНК осталась функция архива, и вроде бы никаких функций ей больше не придумали. У человека примерно 30.000 генов, и набор генов во всех клетках организма одинаковый. Есть исключения, например, эритроциты у млекопитающих, где ядро отсутствует, чтобы им было удобнее переносить кислород, и там просто нет генов. Или половые клетки, в которых при созревании происходит перемешивание генетического материала, и потом он делится пополам. Но это особые случаи. Итак, во всех клетках организма набор генов одинаков, но клетки при этом разные: есть клетки эпителия, есть волосяные луковицы, которые продуцируют волос, есть слизистая оболочка желудка, и т.д. Волосы в желудке не растут, а слизь не выделяется на коже. Почему? Потому что в каждой клетке работает определенный набор генов (слайд 3). Многие гены молчат, и в разных клетках молчат разные гены. Образно выражаясь, можно сказать, что в каждой клетке "звучит" свой аккорд генов, и, как на клавиатуре можно сыграть множество разных мелодий, так и в клетках после оплодотворения зиготы начинают "звучать" разные аккорды, создавая разные мелодии и тем самым уводя клетки по разным путям развития ( слайд 4). Набор генов у разных людей примерно одинаков (слайд 5). У каждого из нас примерно 30.000 генов. Но существуют небольшие различия: некоторые гены у каких-то людей не работают, работа других может заметно отличаться. Иногда эти различия никак не проявляются, но иногда разница является весьма существенной. Например, если не работает всего-навсего один ген, отвечающий за рост организма, возникает заболевание ахондроплазия: нарушение роста костей и ряд других нарушений. Результат показан вот здесь (слайд 6): это картина Веласкеса “Менины”, и любимая карлица инфанты имеет все признаки этого заболевания. Это результат изменения в работе всего одного гена - гена рецептора гормона роста. Те индивидуальные различия, которые отличают нас друг от друга – это результат мутаций, давних или недавних. Мутация – это изменение текста ДНК. ДНК состоит из четырех "букв" - нуклеотидов, которыми записана генетическая программа развития организма. Ошибки при "переписывании" этих "букв" в процессе передачи наследственной информации от клетки к клетке, из поколения в поколение и есть мутации. Некоторые мутации сохраняются тысячелетиями, а некоторые появились только у наших родителей. Чтоб пояснить, как мутации меняют структуру генов, я вспомнила анекдот, когда рабочий приходит наниматься на работу, и его спрашивают: “Что Вы можете делать?” Он отвечает: “Я могу копать”. - “А еще что?" – "Еще могу не копать”. С помощью этой аналогии (слайд 7) я попробую объяснить, как мутации меняют работу генов. Итак, ген может работать, возьмем это за исходный уровень. Мутация может изменить работу гена или изменить уровень его активности, а может привести к выключению гена. При этом выключение гена или изменение уровня его активности можно получить за счет мутаций в структурой части, которые портят белковый продукт, или за счет мутаций в регуляторной части гена, и тогда при хорошей структурной части ген просто не включается и не может сказать свое слово ( слайд 8). Если мутация только немножко испортила белковую последовательность или регуляторную часть, в принципе, можно ожидать, что произойдет обратная мутация, и ген каким-то образом починится и восстановится. Но существуют мутации, когда ген просто теряется. И тогда изменение необратимо. Как и добросовестный рабочий, ген может быть очень активен, и это соответствует мутации, которые увеличивают активность либо белка, кодируемого геном, и он начинает работать быстрее, либо активность регуляторного участка, и тогда синтезируется много копий РНК по сравнению с исходной ситуацией и много белкового продукта. Мы можем нанять много рабочих, чтобы быстро сделать работу, и в ДНК есть аналогичная ситуация, когда создается несколько копий гена. Совсем недавно была опубликована очень интересная статья, в которой говорилось, что люди отличаются друг от друга числом копий гена амилазы. Амилаза - это фермент, расщепляющий крахмал. И оказалось, что народы, которые потребляют крахмалистую пищу, имеют в среднем 7 копий гена амилазы, а те, которые не едят так много крахмалистой еды, имеют всего 5 копий. Комментарий к этой научной работе назывался “ксерокопированный ген”. Если ешь много крахмалистой еды, то выгодно, когда у тебя много фермента амилазы, поэтому в таких группах преимущество имели люди, у которых из-за мутаций число копий гена увеличивалось. Есть еще особый тип мутаций, который воздействует только на регуляторный участок, и эти мутации меняют время работы гена. Ген может работать на разных стадиях развития, долго или совсем краткий период, и в результате мутации меняется время начала или завершения каких-то процессов, например, роста костей. И последний тип мутаций: этот рабочий решил поменять свою специальность и научиться играть на скрипке. Есть мутации, которые меняют специализацию белка, и они уклоняются от своих исходных обязанностей, меняют "профессию". Новая функция – это обычно изменение структурной части гена. Теперь мы знаем, как работают гены, как мутации меняют их, и можем теперь попробовать ответить на вопрос, как обезьяна превратилась в человека. Даем такое техзадание генетику – превратить обезьяну в человека (слайд 9). Что нужно сделать? У обезьяны слишком много волос на теле, нужно выключить ген, который контролирует образование белка волос – кератина, и их станет меньше (слайд 10 и слайд 11). По сравнению с человеком у обезьяны очень длинные конечности. Нужно сократить время работы генов, отвечающих за рост конечностей, и они станут покороче (слайд 12 и слайд 13). Теперь неплохо бы добавить обезьяне мозгов (слайд 14 и слайд 15). Такие гены, которые регулируют размер мозга – не один, а несколько – были обнаружены недавно. Один из них называется ген микроцефалина, его изменения были обнаружены у больных с уменьшенным размером мозга. Потом оказалось, что человек отличается по этому гену от обезьяны. Что еще добавить нашей обезьяне? Наверное, хорошо было бы, чтобы она научилась говорить (слайд 16). Гены, которые контролируют образование структур мозга, необходимых для обучения речи, существуют. Один из них нашли при исследовании семьи, где часто встречались нарушения речи. Этот признак передавался как болезнь с определенным типом наследования, больные члены этой семьи не могли освоить правила грамматики, они не могли научиться правильно говорить, и у них была легкая степень умственной отсталости. У больных нашли мутацию в гене, который называется FOXP2. Потом было показано, что люди отличаются от обезьян по этому гену. При исследовании на мышах выяснили, что он работает во время эмбрионального развития в определенной зоне мозга, регулирует работу других генов, определяя какой из них включается в работу на данном этапе эмбрионального развития. Руководит работой генов, которые участвуют в формировании зон мозга. Гены, которые отличают нас от шимпанзе, которые изменились в процессе эволюции обезьяны, превратившейся в итоге в человека, выявляют, сравнивая ДНК человека и шимпанзе. Вы, наверное, слышали, что существует проект “Геном человека”, и что ДНК человека полностью прочтена. Также сейчас уже прочтена ДНК шимпанзе, и, в большей или меньшей степени, ДНК других приматов. Сравнивая их, генетики пытаются понять, чем мы отличаемся от других приматов и найти те гены, которые отвечают за различия. Их можно назвать "Гены, которые сделали нас людьми". Те гены, накопление мутаций в которых привело к появлению человека. Мутации возникают все время, как песчинки падают в песочных часах. Предполагается, что процесс накопления мутаций идет примерно с одной и той же скоростью, хотя есть исключения. Но многие мутации, возникнув, тут же исчезают. А некоторые остаются и передаются следующему поколению. По последовательности "букв" ДНК человек отличается от шимпанзе примерно одной буквой из ста, при этом друг от друга мы отличаемся одной буквой из тысячи. Это приблизительная оценка. Отличие одной буквы из ста – это изменения, которые появились и в линии шимпанзе, и в линии человека после их разделения. Много это, или мало? Зависит, конечно, от того, где они расположены и что менять, потому что некоторые мутации никак не проявляются (они "нейтральны"), а какие-то являются очень существенным. Сейчас известны некоторые гены, изменения которых привели к появлению человека. Их интенсивно изучают. Поэтому через некоторое время генетики будут знать, какие гены нужно поменять, чтобы сделать из обезьяны человека. У нас есть человек и обезьяна, мы можем их сравнить, и узнать, что это за гены. А как же это происходило 5 млн. лет назад, когда требуемый результат был неизвестен? Остановимся еще на одной детали: что такое мутации и как они возникают? Это изменение "букв"-нуклеотидов в последовательности ДНК.  Светлана Боринская (фото Наташи Четвериковой) Следует различать процесс появления мутации в ДНК и другой процесс – то, что эта мутация не исчезнет сразу, а сохранится хотя бы на протяжении нескольких поколений или распространится и появится у людей в отдаленном будущем. Процесс появления мутации – это химический процесс. Они возникают достаточно случайно, это просто химический процесс, который приводит к изменению молекулы. А распространение мутаций в последующих поколениях у людей (также как у других организмов) – процесс популяционно-генетический. На картинке (слайд 17) показан ряд людей, которые отличаются по какому-то признаку. В следующем поколении соотношение изменится, так как у кого-то не будет детей, у кого-то будет их много. Еще через поколение опять произойдет изменение, и, по случайной причине некоторые признаки могут исчезать, а какие-то становиться всеобщими. Изменение частоты признаков может происходить как случайно, так и целенаправленно, под воздействием отбора. Отбор задается условиями среды, и в разных условиях среды могут отбираться разные признаки (слайд 18). В одних условиях могут быть отобраны одни варианты, например, темная кожа, а в других – светлая. Кожная пигментация – это, генетически определяемый признак, который, наверное, имел наибольшие социальные и политические последствия в истории человечества. Но есть и ряд других признаков, которые не привлекали столь большого внимания общества, как цвет кожи, но, тем не менее, разные по происхождению группы людей могут сильно отличаться по частоте того или иного признака. Легко проиллюстрировать, как среда диктует условия отбора по определенному признаку. При исследовании спортсменов выяснилось, что они отличаются по вариантам гена мышечного белка актинина – это белок, который связан с кислородным обменом в мышцах. При дефиците этого белка усиливается аэробный обмен, а когда его много, то идет анаэробный. Я напомню: если мы устаем, и у нас болят мышцы, то это накопление молочной кислоты, которая не успевает окислиться при недостатке кислорода. Т.е. мышцы работают так интенсивно, что кровь не успевает их обеспечить должным количеством кислорода для окисления появившихся продуктов обмена. А потом, во время отдыха, они окисляются и выводятся, и тогда наши мышцы перестают болеть. Оказалось, что спортсмены, которые занимаются силовыми и спринтерскими видами спорта, где бывает огромная кратковременная нагрузка, когда мышцы работают анаэробно, такие спортсмены имеют генетические отличия от обычных людей (слайд 19). Оказалось, что по сравнению с общей группой, которую использовали в качестве контроля, у этих спортсменов реже встречаются гены "аэробного" белка. Особенно редки среди них были варианты, когда и от мамы, и от папы получены такие "аэробные" варианты гена актинина. А среди олимпийцев вообще ни одного не нашли, чтобы оба имеющихся варианта гена (и тот, который получен от мамы, и тот, что от папы) были "аэробные". Видимо, нельзя стать олимпийцем в силовых видах спорта, если ты получил от кого-то из родителей такой ген. У стайеров, у которых работа идет на выносливость, у них больше частота "аэробного" гена, и у олимпийцев-стайеров она еще больше. Т.е., определенная задача, поставленная перед этими спортсменами, произвела отбор, и если бы этот отбор распространялся бы на потомство, скажем, размножались бы только спринтеры, а стайеры не стали бы размножаться, то мы получили бы иное соотношение частот этих вариантов в потомстве. Признаки потомства бы изменились. Еще один пример, как среда производит отбор. В Юго-Восточной Азии распространен вариант гена, контролирующего окисление этилового спирта, который дает после приема спиртного быстрое накопление ацетальдегида – токсичного продукта окисления алкоголя, того самого, от которого болит голова и возникают прочие неприятные симптомы. Почти у половины населения Юго-Восточной Азии не работает фермент, который обезвреживает это токсичное вещество. А у большинства алкоголиков этот фермент активен. Соотношение частот разных вариантов генов в населении в целом и у алкоголиков показано на рисунке (слайд 20). Видно, что у алкоголиков соотношение изменено. Среди них меньше людей, у которых этот фермент неактивен, просто потому, что при неактивном ферменте накопление токсичного ацетальдегида не позволяет выпить столько алкоголя, чтобы стать алкоголиком. Но это пример отбора, который происходит по сиюминутным запросам среды. А каким образом происходил отбор в эволюции человека? Для рассмотрения этого вопроса я позаимствовала фрагмент из лекции Кирилла Еськова, и я его прочту. "Рамапитек – это один из вариантов «азиатского проекта», который был параллелен африканскому…" - это говорит о том, что одновременно в разных частях света шел процесс гоминизации, т.е., превращения обезьяны в человека." В Азии тоже создавали крупного прямоходящего примата, но на основе не шимпанзе, а орангутана. Там были, например, замечательные гиганты – мегантроп и гигантопитек. И одним из вариантов были рамапитек и сивапитек. И очень даже может быть, что со временем они бы до чего-нибудь даже доэволюционировали. Но в любом случае «африканский проект» успел раньше, и они решили проблемы со всеми, кто им мешал. На этом месте постоянно просятся аналогии, что «проводится тендер». Нескольким конструкторским бюро дается заказ на определенное изделие. Они выставляют его на конкурс, дальше идут стендовые испытания и пр. Потом в итоге одни исчезают, и принимается некая одна модель. Поэтому идея «направленности эволюции», на чем сломано множество копий, обретает на этом месте плоть, на первый взгляд. В эволюционистской картине мира, естественно, заданий никто не дает. Потому что науке категорически противопоказан вопрос «для чего?» ". Конец цитаты. Я попробую рассмотреть, а был ли заказчик в процессе появления человека, а если был, то что именно он заказывал. Здесь на схеме (слайд 21) представлено древо видов, которые также, как и наш вид, шли по пути гоминизации, но не дошли до финала. Справа на схеме Homo erectus, человек прямоходящий, который вышел из Африки и развивался в Азии, а затем вымер. В это время в Африке древо гоминид поветвилось, и около 300 тысяч лет назад одна из ветвей вышла из Африки успешно заселила Ближний Восток и южную часть Европы. Это был неандерталец. Сколько ветвей оставалось в Африке, мы не знаем. Среди них были и наши непосредственные предки. Около 60-70 тысяч лет назад из Африки вышел Homo sapiens, который вытеснил все остальные виды. Некоторое время Homo sapiens и неандерталец существовали параллельно. Чтобы выяснить, были ли заказчик такого направления эволюции, я хочу начать с самого начала: с появления жизни на земле. По современным представлениям (не генетическим), Земля возникла более 4 млрд. лет назад, а жизнь на Земле появилась примерно 3,8 млрд. лет назад (слайд 22). Эта цифра не очень точная, но нам с вами это не важно, а важно только то, что жизнь появилась. Появилась клеточная форма жизни, в виде так называемых прокариотических или безъядерных организмов, из которых потом появились ядерные организмы, которые потом успешно стали многоклеточными, а среди них появились млекопитающие, а среди них – приматы, а из приматов выделились наши предки, и так пока не появились мы и не пришли сюда слушать лекцию. При этом каждый новый уровень сложности возникает на базе предыдущего. Предыдущий при этом никуда не исчезает и исчезнуть не может. Каждый новый вид должен быть вписан с экосистему, в которой он возникает, должен быть адаптирован к тем процессам, к тем источникам пищи, который в этой экосистеме существуют. А пищу на нашей планете и раньше создавали и сейчас создают бактерии, грибы, растения. И если уничтожить бактерий, то рухнет вся система жизнеобеспечения на планете. Никто другой за них их биосферную работу сделать не сможет. Животные потребляют растения, измельчают их при этом, тем самым помогают круговороту веществ в природе, так как окислить неизмельченные крупные растения и вернуть их компоненты в биосферу гораздо сложнее, грибы и бактерии не справляются. А хищные животные собирают, так сказать, сливки с биосферы – они получают концентрированные ресурсы, которые надо только изловчиться поймать. Попутно они поддерживают эти ресурсы в хорошей спортивной форме, чтобы на зубок попадались. Это в очень упрощенном виде нынешнее распределение экологических функций между разными организмами. Похожее распределение существовало всегда. Всегда кто-то кого-то ел, возвращая составляющую еду органику в круговорот веществ. И сам являлся едой для кого-то другого. Шла вполне понятная конкуренция – съесть самому и не быть съеденным другими, оставив при этом потомство. Эволюционные изобретения и уловки для решения этой задачи очень разнообразны. Сегодня нас интересует один из вариантов решения, который оказывался актуальным, когда другие, более простые решения, не срабатывали. Это получение преимуществ за счет кооперации усилий и разделения обязанностей. Например, симбиоз. Объединяются организмы, каждый делает что-то полезное, и в определенных условиях у них больше шансов выжить вместе, чем по отдельности. Путем симбиоза из безъядерных (прокариотических) организмов появились более сложные ядерные (эукариоты). Но есть и другой способ повышения эффективности питания и защиты – это кооперация с себе подобными. Переход от одноклеточных к многоклеточным. Затем, на другом уровне, кооперация многоклеточных с образованием социальных систем, таких как социальные насекомые (пчелы, термиты) или социальные млекопитающие. Вот следующий шаг - объединение простых социальных систем в более сложные, а затем сложных в суперсложные – сделал только человек. Я дам очень краткий обзор дальнейшей социальной эволюции, т.к. на первой лекции Кирилла Еськова был вопрос, в чем применимы представления биологической эволюции к социальной. В конце лекции даны ссылки на работы специалистов по этой теме. Полагаю, что здесь должна идти речь не о применимости представлений о биологической эволюции к социальной, а о том, что эволюция и биологических систем, и социальных имеет общие закономерности, свойственные эволюции систем вообще. Самая простая стадия социальной эволюции человека – это община. Общины бывают эгалитарные, где все равны, или неэгалитарные, где у кого-то ресурсов и власти больше, у кого-то меньше. Численность общины бродячих охотников-собирателей – у культурных антропологов она называется “ЛОКАЛЬНАЯ ГРУППА” - не превышает двухсот человек, а обычно равна 20-50 чел., крайне редко в каких-то очень плодородных районах может достигать 500 человек (в случае если речь идет об оседлых высших охотниках-собирателях или земледельцах). При образе жизни охотников-собирателей плотность населения составляет примерно один человек на 10 кв. км. (по крайне мере у тех, которых успели изучить в XIX-XX вв.) Охота-собирательство – это тот образ жизни, который существовал не только сотни тысяч лет человеческой эволюции, но до этого миллионы лет существовал у обезьян. Если в социальной системе больше 200 человек, то организация должна быть уже более сложной. Более сложный уровень – это объединение нескольких общин, называемое ВОЖДЕСТВОМ. Объединение происходит чаше всего через завоевание, при этом одна из общин становится главной, а остальные ей подчиняются. Альтернативой такой системе является эгалитарная система, когда несколько общин объединяются в единый социальный организм, но главная община не выделяется. Еще более сложное общество – это когда образуется система вождеств, или за счет завоевания одного вождества другим, или за счет деления разросшегося вождества. Численность населения СЛОЖНЫХ ВОЖДЕСТВ от 5 до 30 тыс. человек, обычно на этом уровне сложности общество уже перешло к земледелию или скотоводству. Альтернативой сложному вождеству является ПЛЕМЯ (этот термин имеет несколько значений, одно из них используют для обозначения обществ с определенной структурой). Племенные объединения могут объединять группы, которые уже прошли через стадию вождества, а могут образовываться и сами по себе путем объединения обществ других видов. СУПЕРСЛОЖНОЕ ВОЖДЕСТВО – объединение нескольких сложных вождеств. Полисная организация, которая имеет сложную историю возникновения, возможно, - результат деирархизации сложных вождеств. Дальнейшее усложнение приводит к появлению ГОСУДАРСТВ, которые образуются либо из сложных вождеств, либо из полисных объединений. Из племен государства не образуются, потому что племя не иерархизировано. Дарвиновский отбор наиболее удачных сочетаний из наследуемых случайных вариаций признаков (здесь признаки – определенные характеристики общин, вождеств и т.п.) применим и к социальной эволюции, но характер варьирования признаков, сами признаки и способ их наследования там, естественно, другой. В рассмотренной выше схеме есть маленькое дополнение к теории эволюции в том виде, как ее излагают во многих учебниках: выделение определенных уровней сложности систем. То есть мы сейчас рассматривали не любые изменения и направления эволюции, а те, в которых происходило объединение элементов в более сложную систему. В систему другого уровня сложности. Самое простое – это безъядерные прокариоты, затем появились более сложные одноклеточные ядерные организмы (как система тесно взаимодействующих прокариот), потом многоклеточные организмы (как система взаимодействующих клеток), затем социальные организмы (системы взаимодействующих многоклеточных), в рамках которых возникло человеческое общество, а далее шли этапы социальной эволюции, характерные только для человека. Такие уровни в эволюционном развитии (слайд 23) были названы Валентином Федоровичем Турчиным метасистемами, а усложнение с переходом на следующий уровень было названо метасистемным переходом. С моей точки зрения, это очень важное дополнение к теории Дарвина во всех ее современных ипостасях. Если затребовано усложнение, т.е., переход на некоторый другой уровень сложности, то этот переход может появиться одновременно в разных местах и тот, кто преобразовался первым, получает преимущество перед остальными и может их победить в конкурентной борьбе (как это показано на слайде 23 в правой части). Как раз тот тендер, о котором говорил Кирилл Еськов. Я это все рассказываю к тому, чтобы подчеркнуть, что всякий вид, который появляется на Земле, должен быть адаптирован к условиям, в которых он появился. Например, когда возникли эукариоты, они возникли не в безвоздушном пространстве, а на Земле, населенной прокариотами. Затем возникли многоклеточные, которые должны были приспособиться к жизни среди одноклеточных. Появление социальных организмов произошло в очень сложной среде, включающей существа и системы всех предыдущих уровней. И, наконец, появился человек, а это произошло в условиях, когда существовали группы других приматов на протяжении миллионов лет. И здесь можно искать тот запрос, который среда предъявила этим приматам (слайд 24). То, что исчезла шерсть, ясное дело, не главное, и прямохождение, хотя и очень полезно, тоже не самый важный принцип. А что было важным? Последний раз редактировалось Chugunka10; 21.11.2021 в 14:37. 
|
|
#2
12.09.2013, 20:34
|
||||
|
||||
|
Реплика из зала. Мышление.
Боринская. А какие преимущества оно дает? Какие функции выполняет мышление? Реплика из зала. Опыт. Боринская. Т.е., передача опыта из поколение в поколение. Она есть уже и у обезьян, и человек тоже мог это делать более эффективно. В передаче опыта принципиальным было появление млекопитающих. У них два поколения в обязательном порядке контактируют, так как мать вскармливает детеныша. Это создает основу для негенетической передачи информации из поколения в поколение, т.е. для появления культуры. Этот способ возник гораздо раньше, чем появился человек. Еще недавно считалось, что у животных не может быть культуры, а теперь идут дискуссии о том, как называть обучение от взрослых, которое есть у детенышей обезьян: называть это протокультурой или культурой, и где граница, которая отличает по этому критерию человека от обезьяны. Итак, мышление позволяет передавать опыт от поколения к поколению, накапливать его. Есть ли еще что-то, для чего пригодилось бы мышление? Реплика из зала. Коммуникация. Боринская. А зачем она нужна? Реплика из зала. Чтобы организоваться с другими людьми, чтобы делать какое-то общее дело. Боринская. Например, успешно добыть еду или от кого-то защищаться. Стали умнее, чем соседнее стадо, мы быстрее добегаем до бананового дерева и лучше обираем бананы. Можно дальше не усложняться. А все-таки дальнейшее развитие происходило. Какие есть идеи? Почему человек на протяжении миллионов лет становился все умнее, все способнее к коммуникации и все организованнее. Реплика из зала. Появилась речь, язык. Боринская. Ну и что, что появилась речь? Дальше усложняться не нужно. Сиди себе и ешь бананы, всех обезьян уже отогнал. Реплика из зала. Конкуренция. Боринская. Замечательная идея: но конкуренция не между индивидами внутри группы, а конкуренция самой группы с другими группами, близкими к эволюционирующей по уровню развития, должна была все время двигать человека к поумнению. Есть и другие теории вроде того, что нужно было какую-то косточку разгрызать или что-нибудь откуда-нибудь выковыривать орудиями, но такие запросы одномоментны, они не могут действовать все время. Т. е., если предположить, что запросом среды на поумнение, усиление развития средств коммуникации, было наличие других групп с аналогичными умениями, и выигрывал тот, кто лучше кооперировался, был лучше организован в защите и нападении, то можно объяснить, зачем человеку надо было неуклонно умнеть, развивать речь и навыки коммуникации. А как мы видели на примере со спортсменами или с алкоголиками, если есть запрос, то гены его обеспечивают. Могу привести интересный пример, как быстро можно развить коммуникацию и социальные взаимодействия. Некоторый косвенный ответ на этот вопрос дают эксперименты, проведенные в Новосибирске по разведению серебристых лис. Их разводили на фермах. У этих зверей очень хороший мех, но сами звери очень агрессивные, их трудно было содержать, поэтому был начат отбор на одомашнивание серебристых лис. Нужно было получить менее агрессивных животных. Это удалось: на расплод отбирали тех животных, которые меньше боялись человека. Их отбирали так: человек приближался к животным, и отмечал, какие лисы меньше его боятся, меньше принимают агрессивные позы, не скалят зубы и т.д. Их и брали на расплод. За двадцать поколений такого отбора удалось получать животных совершенно одомашненных (слайд 25). Появились лисы, которые, как собачки, ластились к человеку, очень тепло к нему относились и очень радовались его появлению. Но кроме этого у них произошли еще какие-какие изменения. У них хвост стал завиваться колечком, тоже как у собачки, опустились уши и они начали тявкать. И еще шерсть у них стала плохая. Хотя по этим признакам отбор не велся. Глазков. А как шерсть была связана с агрессивностью или изменением среды? Боринская. Оказалось, что это сопряженные изменения. У этих лис сейчас выявлены еще не все гены, но некоторые гены, которые изменились в результате такого отбора, уже известны. Это гены, контролирующие работу гормонов и регулирующие передачу нервного импульса. Дело в том, что некоторые вещества, работающие в нервной системе при передаче импульса, участвуют и в общем обмене, и в том числе в выработке пигментов. Есть общие звенья путей синтеза веществ, участвующих и в образовании пигментов, и в передаче сигналов. За счет того, что шел отбор по одному признаку, был зацеплен и другой. Глазков. Это просто случайность или это зависит функциональным образом от поведения? Боринская. Это особенность структуры обмена веществ. Я могу привести пример, когда антропологи пытались выяснить, отличаются ли больные шизофренией от здоровых, провели кучу измерений и в конце концов выявили один признак: у больных были более темные волосы. Это не значит, что все брюнеты – шизофреники. Дело в том, что при шизофрении меняется обмен вещества, которое называется дофамин, а темный пигмент образуется из того же предшественника, что и дофамин. Поэтому при изменении синтеза дофамина захватываются и какие-то звенья синтеза пигмента. Глазков. Т.е., в данном случае, с лисами это случайность? Они теоретически могли стать и красивее. Боринская. Мы оставим это на потом, и я расскажу, зачем вообще все на Земле эволюционирует. Жили бы бактерии, не менялись, чем им плохо? Почему все не остановилось на каком-то уровне развития? Видимо, есть непрерывный запрос среды. Вот здесь на картинке, которую я взяла из статьи Георгия Александровича Заварзина, интересный аспект устройства природы – круговорот веществ (слайд 26). Биогеохимическая машина планеты. И она, эта машина, контролируется живыми организмами, т.е., на каждом цикле и на любом доступном процессе сидят какие-то виды бактерий, которые этот процесс контролируют. На них сидят более организованные существа и их едят и контролируют их численность, и т.д. Эта система создает запросы на появление новых видов и на создание новых экологических ниш. Я предполагаю, что именно эта система была заказчиком эволюции человека. А эволюция эта протекала в тех условиях, которые тогда существовали. А существовали на тот момент группы животных, высокоразвитых приматов. И здесь есть еще один важный момент, негенетический – как человек и его цивилизация взаимодействуют с биосферой. Существуют две модели взаимодействия: одна – это то, что цивилизация – это некий спрут, который сидит на биосфере, ее выжирает, и скоро все рухнет. А другая – то, что цивилизация вписана в биосферу, что существуют некоторые механизмы регулирования аппетитов человечества, и существование таких механизмов защищает биосферу - по крайне мере частично - от разрушения человеком, и самого человека от полного подрыва своей биосферной базы. Эта идея проиллюстрирована картинками (слайд 27), которые я взяла из презентации географа Дмитрия Люри. Очень интересны механизмы, которые регулируют деятельность человечества. По мере исчерпания ресурсов человеку приходится менять способы добычи ресурсов, и, соответственно, среду своего существования. И вот тут мы снова можем вернуться к генам. То, как сам человек меняет среду своего обитания по мере развития цивилизации, отражается в его генах. Две коротких иллюстрации того, что происходило с человеком после того, как он появился. Еще раз напомню, что он появился как охотник-собиратель, это группы, кочующие по определенной территории, вылавливающие там животных и собирающие съедобные растения. Раньше, 15 тысяч лет назад, все люди были охотниками-собирателями. 500 лет назад охотников собирателей остался только один процент, а к концу XX века таких групп осталось очень немного, а все остальные люди перешли к цивилизованному образу жизни: земледелию, скотоводству и другим занятиям. При этом окружающую среду можно разделить на компоненты: природный (климатические условия, ландшафт), биогенный (доступность пищи, наличие инфекций) и антропогенный – это та часть среды, которую человек создал сам. Рассмотрим примеры генетических изменений, вызванных этими разными факторами. Адаптация к климатическим факторам. Еще в XIX в. было замечено, что, если есть группы, которые различаются величиной, то более мелкие виды тяготеют к более теплому климату, а более крупные – к холодному, т.к., если тела крупные, то потеря тепла меньше. Это верно и для человека. Антропологи уже давно определили, что средний вес людей в разных климатических и температурных зонах разный (слайд 28). Еще одна закономерность, найденная для – выступающие части тела длиннее там, где тепло, короче там, где холодно (слайд 29). Такие же примеры различий в строении тела можно найти и у людей (слайд 30). Эти различия наследуются, то есть определяются генетически. Устойчивость к инфекциям. Известно, что в районах, где водится малярия, есть и защитные мутации организма к ним. Интересно, что эти мутации были обнаружены во время корейской войны, когда американским солдатам давали лекарство примахин, которое должно было уберечь их от малярии. Но некоторые солдаты умерли не от малярии, а от самого этого лекарства. Когда провели расследование, выяснилось, что эти солдаты несли мутацию в одном из генов, и происхождением они были из Средиземноморья или из Африки, т.е., из районов, где распространена малярия. Так была открыта одна мутация, потом оказалось, что их несколько. Они возникли независимо, но обеспечивали решение одной и той же задачи – устойчивость к малярии (слайд 31). Аналогичный пример связан не со средой, а с появлением новой пищи. При одомашнивании животных человек получил доступ к молоку. У всех млекопитающих детеныш во время вскармливания может переваривать молоко, а по окончании этого периода фермент, который расщепляет молочных сахар, исчезает. Но у некоторых людей синтез этого фермента продолжается до взрослого состояния. А если этого фермента нет, то можно выпить молока, но потом будет очень плохо: живот будет болеть, и т.д. Оказалось, что у финнов 100% младенцев усваивает молоко, а из взрослых только 85% имеет этот фермент и может пить молоко, а у 15% взрослых фермента нет и от молока им плохо. А у японцев только 2% взрослого населения может пить молоко без последствий (слайд 32). Мутация произошла в регуляторном участке гена, который кодирует фермент, расщепляющий молочный сахар, и этих мутация несколько: одна из них распространена в Европе, вторая на Ближнем Востоке, третья в Африке (слайд 33). В Азии пока такие мутации не открыты, и население там, в основном, молоко не пьет. Процесс отбора мутаций был тот же самый: был дан заказ от среды, и гены его реализовали. И последняя иллюстрация того, как может действовать отбор. Здесь (слайд 34) представлены данные об изменении численности населения в Китае в течение последних двух тысяч лет. Китайские императоры проводили перепись населения, не для того, чтобы нам было удобно это исследовать, а с целью налогообложения. Когда эти данные были собраны вместе, оказалось, что каждые 150-300 лет происходили резкие падения численности, вот здесь, например, с 60 млн. до менее чем 20-ти. Такие кризисы происходили не только в Китае, но и в других регионах. После введения современной медицины они стали не столь резкими. Во время кризисов действуют несколько факторов: война, голод и эпидемии. Если это голод или эпидемии, то ясно, что кризисы будут действовать как мощный фактор отбора тех вариантов генов, которые будут защищать людей от эпидемии или делать их более устойчивыми к перебоям с пищей. Периодические колебания численности ускоряют действие отбора и сопровождаются изменением важных для выживания генетических характеристик. Я хочу поблагодарить тех, дискуссии с кем мне помогли подготовить этот доклад - Николая Янковского, Владимира Спиридонова и Владимира Алешина. Спасибо всем за внимание. Обсуждение Долгин. Мы сейчас устроим очень маленькую дискуссию, потому что очень перебрали время, поэтому все остальные вопросы мы соберем и вывесим на сайте и попросим Светлану ответить на них на сайте, а сейчас очень коротко, в режиме блица. Вопрос из зала. Два вопроса: считаете ли вы, что ваша теория совместима с теорией высшего начала или вы все объясняете без обращения к нему? И второе: с вашей точки зрения, гомо сапиенс может эволюционировать и можно ли просчитать время его существования до того, как он превратится во что-нибудь другое? Светлана Боринская (фото Наташи Четвериковой) Светлана Боринская (фото Наташи Четвериковой) Боринская. Эта теория абсолютно совместима с наличием высшего начала, поскольку высшее начало ни разу не нарушало эти правила. Долгин. Теория не противоречит, но и не нуждается в этом начале. Правильно я понимаю ваш ответ? Боринская. Да, совершенно верно. Пока что здесь не было ни одного аргумента, где потребовалось бы стороннее вмешательство. Это была игра по правилам. Вопрос из зала. А кто придумал правила? Боринская. На этот вопрос я не смогу сейчас ответить, это за пределами генетики. Насчет эволюции человека: эволюцией считается образование видов – макроэволюция, и микроэволюция – эволюция внутри вида. Отбор все время происходит, как внутри одного вида, так и между ними, просто их существует два вида: движущий и стабилизирующий. На нас с вами, естественно, отбор действует – не все ныне живущие люди оставят потомков. Если среда создаст запрос, с которым мы не сможем справиться в рамках нашей генетической и социокультурной адаптации, то придется тогда эволюционировать в новый вид. Лев Московкин. Не было сказано ни слова ни про митохондрии, ни про то, что, как мне кажется, является вашим коньком – <неразборчиво> на социальное поле, точно так же как Гельфанд например, ничего не сказал про сплайсинг. В этом и состоит вопрос. Боринская. О "митохондриальной Еве" и других аспектах изучения генетиками эволюционной истории человека можно прочесть здесь. Вопрос из зала. Я поддерживаю то, что жалко, что не было сказано про митохондриальную Еву, но вопрос у меня про клавиатуру, из которой природа извлекает процесс эволюции, извлекает аккорды, которые и звучат. Исторический процесс формирования этого генома для меня остался за кадром вашего выступления, и я не знаю, что про это можно сказать. Когда он такой огромный и замечательный, из которого можно было извлекать уже практически все, добавляя к нему только чуть-чуть, в какой период времени он формировался, и это было плавно или скачками? Боринская. По размеру генома человек от обезьяны не отличается. Геномы всех млекопитающих достаточно близки по составу, есть отличия от более отдаленных родственников, но тогда надо было бы сделать об этом отдельный цикл лекций. Вопрос из зала. Но где разрывы в эволюции генома? Он менялся по эволюционой лестнице плавно или скачками? Боринская. Отдельная мутация появляется скачком - одномоментно, в генеративных клетках родителей, и, чтобы выйти на эволюционную арену, она должна попасть к потомку. Если мутация не отсеяна отбором на ранних стадиях развития этого конкретного потомка, то он получает возможность передать ее своим детям. Но с точки зрения эволюции пока ничего еще не произошло – даже если эта мутация сильно меняет какой-то признак, то частота ее все равно ничтожна, одна особь из сотен тысяч или миллионов. Чтобы возник новый вид или хотя бы новый признак, заметный на популяционном уровне (т.е. изменение, которое можно будет увидеть в палеонтологической летописи), частота мутации (и соответствующего признака) должна возрасти. Скорость возрастания частоты определяется, во-первых, числом потомков, которых особь может произвести, во-вторых, интенсивностью отбора, то есть тем, насколько быстро подохнут потомки других особей, этой мутации не имеющие. Допустим, на популяцию напал мор, и численность ее сократилась на порядок. Если носители нашей мутации выживают более успешно, (устойчивы к инфекции, например, или могут переваривать какую-то необычную, недоступную другим пищу, что окажется принципиальным для выживания), то после мора их станет в 10 раз больше, например, из 1% получится 10%. Они станут заметны на популяционном уровне. Тогда мы сможем сказать "возник новый признак у части особей". Если повторить мор несколько раз, то их частота возрастет еще больше, или они даже вытеснят всех не имеющих мутации. Тогда мы сможем сказать "признак изменился у всех". Эта последовательность событий не похожа на скачок. То есть это такой скачок, который включает несколько этапов. А если учесть, что мор не каждый эволюционный день бывает, и что большинство признаков определяется не одной мутацией, а многими (особенно когда речь идет о полезных признаках, с вредными проще, ломать – не строить, испортить дело можно и одной мутацией), то "скачки", можно сказать, становятся такими плавными и, при мелкомасштабном рассмотрении, ненаправленными шагами по эволюционному пространству. Это пространство – не лестница, лестница возникает при рассмотрении только одного направления многомерного пространства эволюционных возможностей, и по ней карабкаются только те виды, которым не удалось втиснуться в возможности адаптации на предыдущих ступеньках. Это отдельный вопрос, я попыталась здесь коротко ответить. Я думаю, что резкость скачков зависит от масштаба рассмотрения вопроса – что считать резким. Вопрос из зала. Два вопроса: почему мир ДНК оказался более эффективным, чем РНК, и почему эволюция ограничилась двумя нитями сначала РНК, потом ДНК и не перешла на тройную, четверную, и т.д., полинить. Это первый вопрос. А второй: о том, что было в первой лекции, когда была выдан тип заданий на то, что мы сейчас называем человеком, были исходные модели, в том числе, и орангутана. И модель орангутана проиграла конкурентную борьбу той модели, что существует сегодня. А встретиться они должны были, судя по всему, сколько-то десятков тысяч лет назад. До какой стадии дошла модель орангутана и что она из себя представляла в момент своего уничтожения? Боринская. Спасибо вам за очень интересные вопросы. Первая половина вопроса была к Гельфанду, а вторая – к Еськову. Долгин. Мы вот-вот повесим возможность задавать вопросы Кириллу Еськову, так что все будет. Вопрос из зала. Добрый вечер. Не секрет, что мир долго верил в существование множества богов. И долго религия говорила, что человек произошел, скажем, от Адама и Евы, а наука – теория Дарвина и т.д. - утверждала что от обезьяны. И сейчас появилась теория на уровень выше о том, что религия и наука – это все одно общее, теория энергетики и т.д. Как генетика может объяснить этот замысел? Потому что сейчас говорится, что все было создано по какому-то замыслу, и одновременно это наука. Долгин. Все-таки не надо путать науку с не наукой. Боринская. Спасибо за очень важный вопрос. Несколько лет назад в одном из ведущих журналов, по-моему, “Nature” или “ Science” была заметка, которая называлась “Ген религиозности”. Но это нужно воспринимать скорее как шутку  . Религия в генах не записана. Религия отражает особенности нашего мышления, это колыбель нашего мышления, а наука существует намного меньше времени, чем религия. И наше мышление, особенно массовое мышление, имеет много особенностей, которые существуют давным-давно. И никакого противоречия в том, что у людей существуют разные картины мира, нет. Одни воспринимают мир таким способом и требуют доказательств, а другие – другим способом и хотят верить без доказательств, либо система доказательств у них построена совершенно по-другому. Для того, чтобы исследовать соотношение разных картин мира, у нас есть очень интересный, проект, который рассматривает географическое распределение древних мифов, историю их расселения, и очень интересно сопоставление истории распространения мифов с миграциями народов. Оказывается, что определенные сюжеты в мифах существуют более 15 тысяч лет, а некоторые, видимо, более 30 тысяч лет. Возможно, эти сюжеты вышли из Африки, а это произошло около 70 лет назад. Так что некоторые мифологемы очень устойчивы и, видимо, это связано с закономерностями социальной эволюции и некоторыми особенностями мышления человека. . Религия в генах не записана. Религия отражает особенности нашего мышления, это колыбель нашего мышления, а наука существует намного меньше времени, чем религия. И наше мышление, особенно массовое мышление, имеет много особенностей, которые существуют давным-давно. И никакого противоречия в том, что у людей существуют разные картины мира, нет. Одни воспринимают мир таким способом и требуют доказательств, а другие – другим способом и хотят верить без доказательств, либо система доказательств у них построена совершенно по-другому. Для того, чтобы исследовать соотношение разных картин мира, у нас есть очень интересный, проект, который рассматривает географическое распределение древних мифов, историю их расселения, и очень интересно сопоставление истории распространения мифов с миграциями народов. Оказывается, что определенные сюжеты в мифах существуют более 15 тысяч лет, а некоторые, видимо, более 30 тысяч лет. Возможно, эти сюжеты вышли из Африки, а это произошло около 70 лет назад. Так что некоторые мифологемы очень устойчивы и, видимо, это связано с закономерностями социальной эволюции и некоторыми особенностями мышления человека.Вопрос из зала. То есть, генетика является определенной картиной мира, так же, как религия, и т.д.? Боринская. Наука – это принципиально другой подход к познанию устройства мира, возникший относительно недавно. Естественные науки, о гуманитарных я сказать не могу, в своем восприятии мира и построении своих тезисов отличаются от религиозных доктрин. Научное исследование не должно иметь логических противоречий. Ученые – это команда людей, которые играют по определенным правилам. Они требуют доказательств не чудесами, а экспериментальными исследованиями. От религиозных трактатов логическая непротиворечивость или экспериментальные подтверждения предложенных откровений не требуются. Способность к логическим рассуждениям не является общераспространенной. Школьный аттестат ее не гарантирует. Ее не гарантирует даже диплом о высшем образовании. Если вы не различаете логически противоречивые и непротиворечивые суждения, то для вас нет разницы между наукой и не-наукой. Люди верят в то, что им удобно, что соответствует их картине мира (которая может быть логичной в одних частях, обычно в области практического опыта индивида, и совершенно нелогичной в других). И они воспринимают на веру утверждения журналистов, потому что броско сказано. Тогда наука, эзотерика, гадания, предсказания астрологов, религия – все в один ряд. И у них никаких сомнений не возникает, что что-то там чему-то противоречит. Но у этих картин мира совершенно разные функции, и смешивать их бессмысленно. Кстати, боги прошлого давали очень полезные рекомендации – когда сеять, когда урожай собирать, когда воевать, какие экологические стратегии осуществлять. Собственно, те боги, которые давали неполезные рекомендации, не могли обеспечить выживание своих адептов и вместе с ними исчезли. Сейчас технические возможности человечества возросли настолько, что принимать решения о налоговой политике, о введении генотерапии или об экологических стратегиях, которые будут иметь глобальные последствия, на основе предсказаний оракула невозможно. Надо обладать профессиональными знаниями. Решения, требующие научных знаний, не могут подменяться религиозными предписаниями. А вера не нуждается в подкреплении научными знаниями, иначе это не вера. Наука и религия – это разные сферы человеческой деятельности. Это никак не мешает некоторым ученым, в том числе и генетикам, быть верующими людьми. Например, когда Жак Моно открыл схему регуляции активности бактериального оперона, он решил, что это доказательство бытия Божьего, потому что как же иначе она получилась бы такая замечательная и стройная. Это была просто его личная точка зрения, что никак не мешало ему делать высококлассные работы в области молекулярной биологии. Долгин. Но при этом не надо воспринимать молекулярно-биологическую картину мира как рядоположенную религиозной. Они в совершенно разных плоскостях. Вопрос из зала. Нет, это не так. Есть общие точки. Долгин. Нет общих точек – они отвечают на разные вопросы. Но есть подходы внутри религиозных систем, которые не мешают заниматься наукой, а есть мешающие этому. Материалы по теме лекции Проблемы эволюции. Сайт Александра Маркова http://www.macroevolution.narod.ru/ Клиодинамика. Сайт Петра Турчина http://www.eeb.uconn.edu/people/turchin/Clio.htm Боринская С.А., Карташова О.В. Системный подход к изучению эволюции ("Биология". 2000. №23.). Идеи и практические советы по преподаванию теории эволюции в школе. Боринская С.А., Янковский Н.К. Люди и их гены. М: Век-2, 2006. 64 с. Тейяр де Шарден П. Феномен человека. М.: Прогресс, 1965. Турчин В.Ф. Феномен науки: Кибернетический подход к эволюции. 2-е изд. М.: ЭТС, 2000. 368 с. Альтернативные пути к цивилизации / Под ред. Н.Н. Крадина, А.В. Коротаева, Д.М. Бондаренко, В.А. Лынши. М.: Логос, 2000. 368 с.: Красилов В.А. Метаэкология.
|
|
#3
22.03.2014, 11:22
|
||||
|
||||
|
http://pereformat.ru/2013/11/nashi-predki/
Опубликовано 21.11.2013 Происхождение человека – одна из наиболее интригующих и захватывающих тем науки, философии, мировоззрения. И одна из наиболее запутанных. Дело в том, что нет ни одного прямого эксперимента, который твердо и однозначно дал бы ответ на вопрос, в каком месте планеты и когда впервые появился наш прямой предок, который подпадал бы под антропологическое описание вида Homo sapiens и/или «анатомически современный человек» (АСЧ). Здесь каждое понятие не зафиксировано и является по сути «плавающим». Нашли древние скелетные остатки, но как узнать, это «впервые», или завтра найдут еще древнее? Насколько надежны датировки, которые на самом деле вовсе не надежны и практически всегда оспариваются? Есть десятки антропологических признаков, которые так или иначе примеряют на понятие Homo sapiens и на понятие «анатомически современный человек», но теория это одно (хотя общепризнанной полной классификации все равно нет), а на практике эти признаки применить в полной мере практически невозможно – обычно находят лишь фрагменты скелета, часто без лицевых костей, и для наиболее древних костных остатков практически всегда наблюдаются некие «архаичные» признаки.  И далее в дело вступает то, что называют добросовестностью научного работника. Ставки велики – каждый новый скелет или его фрагмент, который дает возможность провозгласить его «древнейшим из известных» Homo sapiens или АСЧ становится всемирной сенсацией, со всеми вытекающими последствиями в виде научных наград, крупных финансовых грантов, выборами в престижные академии наук. Поэтому, к сожалению, так нередки искажения данных, описываемых в академической и прочей печати, не говоря о популярных изданиях, так охочих на сенсации. Датировки в научных изданиях порой завышаются, архаические признаки «замазываются», и становится совсем трудно разобраться, где настоящие данные, и где фантазии авторов. Нужны перекрестные исследования, которые редки. Наконец, немало просто неквалифицированных работ, особенно в области популяционной генетики, или работ, ориентированных на заранее заданный результат. Об этом и будет наше повествование. А именно, о том, как трудно пробить стену ангажированных исследований, которые «заточены» на якобы выход «анатомически современного человека» якобы из Африки, и что на самом деле показывают исследования, часто тех же авторов, но своеобразно ими интерпретируются. Стена к тому же цементируется идеологическими соображениями, по которым надо любой ценой показать «африканское происхождение человека», а кто находит другие данные и проводит другие интерпретации, тот «расист». Стена укрепляется и тем, что практически все статьи ангажированных исследователей, а это большинство популяционных генетиков, начинаются фразой «Как известно, анатомически современный человек вышел из Африки». То есть установка идет с самого начала. Это резко повышает вероятность публикации статьи в академическом издании. Вот несколько примеров, взятых из названий академических статей, или из первых фраз вступлений в статьи: Происхождение человека: выход из Африки (название статьи; Tattersal , 2009); Эволюция человека и выход из Африки (из названия статьи; Stewart and Stringer, 2012); Африканское происхождение мужского (генетического) разнообразия (из названия статьи; Cruciani et al, 2011); Африканское происхождение современных людей Восточной Азии (из названия статьи; Ke et al, 2001); …анатомически современные люди прибыли в Европу из Африки по меньшей мере 45 тысяч лет назад, вслед за распространением людей из Африки (Moorjani et al, 2011); Полагают, что современные люди произошли в восточной Африке (Henn et al, 2011); Общепризнано, что анатомически современные люди произошли в Африке (Hammer et al, 2011); Африка, прародина всех современных людей (Lachance et al, 2012); …расхождение анатомически современных людей из Африки было примерно 44 тысячи лет назад (Underhill et al, 2000); Современные люди произошли в Африке примерно 200 тысяч лет назад (Campbell and Tishkoff, 2010); …Анатомически современные люди произошли из небольшой изолированной популяции в Африке 150-200 тысяч лет назад (Patin et al, 2009); Суб-Сахара и северо-восточная Африка – наиболее вероятные регионы происхождения человека и коридор в остальной мир (Arredi et al, 2004); …расхождение человека началось в Африке (Ramachandran et al., 2005). Ниже в этой работе будет показано, что все эти положения, и аналогичные им, которые вопроизводятся в десятках и сотнях академических и прочих статей – неверны. Вопрос – как наука о происхождении человека дошла до жизни такой? Как мог сложиться «консенсус специалистов», основанный на однобокой и накатанной интепретации под заранее заданный ответ? Как вообще в науке могла сложиться ситуация, когда другие обоснованные интерпретации тех же или других данных встречаются выраженной агрессией, политическими обвинениями, неприкрытыми негативными эмоциями? Почему «выход из Африки» стал религией, основанной на вере, которая в доказательствах не нуждается? До 1980-х годов обсуждения африканского происхождения человека шли вяло, и были по сути маргинальными. Рассматривать это серьезно мешали два обстоятельства. Первое – было признано, что далеким предком современного человека был Homo erectus, человек прямоходящий, который появился несколько миллионов лет назад, возможно, в Африке, но было известно, что уже почти два миллиона лет назад он распространился по всей Евразии. Поэтому Homo sapiens, человек разумный, мог стать его потомком где угодно. Второе – было показано, что ближайший родственник АСЧ, неандерталец, в Африке не жил. Поэтому общий предок современного человека и неандертальца, который жил по разным данным между 600 и 300 тысяч лет назад, получается, что в Африке тоже не жил. К тому же у неандертальца была светлая кожа, и на этом мы остановимся ниже. Поэтому африканское происхождение современного человека требует прибытия светлокожего непосредственного предка человека в Африку, скажем, 500-300 тысяч лет назад, далее было самостоятельное, эволюционное приобретение им черной кожи, иначе в Африке не выжить, а потом выход его из Африки и обратное самостоятельное превращение черной кожи в светлую. На этот счет была даже придумана остроумная гипотеза о роли витамина D в самостоятельном (без скрещивания со светлокожими людьми, которых быть за пределами Африки не могло, иначе концепция рушится) превращении чернокожих в светлокожих, но эта гипотеза никогда не была подтверждена экспериментально. Так и осталась умозрительной. В общем, до середины 1980-х об африканском происхождении современного человека говорить было не очень серьезно. Но потребность в этом «в определенных научных кругах» или, точнее, среди ученых определенного либерального мировоззрения явно назревала, иначе не объяснить последующее развитие событий. А произошло то, что в 1987 году в журнале Nature была опубликована статья Ребекки Канн с соавторами из Калифорнийского университета, Беркли, под названием «Митохондриальная ДНК и эволюция человека». Статья весьма слабая не только по современным критериям, но и по тогдашним, и можно только удивляться, как статья прошла рецензии. Достаточно упомянуть, что в Абстракте, предваряющем статью, сообщалось, что изученные авторами митохондриальные ДНК происходят от одной женщины, которая, «как было постулировано» (!), жила примерно 200 тысяч лет назад, «предположительно» (!) в Африке. После публикации статьи разверзлись хляби небесные, распахнулись шлюзы и ворота. Восторг западной прессы о том, что африканцы – наши предки, был фантастическим. Эту древнюю африканскую женщину тут же окрестили Евой, и ведущие журналы мира поместили информацию об этом на глянцевые обложки. С тех пор настойчивая обработка общественного мнения продолжается без остановки, если не по нарастающей. Это стало общепринятым мнением, оспаривать которое сродни утверждению о возможности вечного двигателя. Иначе говоря, оспаривающий идет против научного «консенсуса», которого, конечно, нет, но который постоянно провозглашается. Обсуждая эту проблему с антропологами, я (как главный редактор международного журнала «Advances in Anthropology») получал и продолжаю получать немало писем, в которых профессиональные ученые делятся тем, что они, разумеется, сомневаются или категорически не согласны, что «африканское происхождение человека» хоть как-то обосновано, но не хотят об этом выступать в печати, потому что «себе дороже». И потому что статью в научный журнал все равно отклонят независимо от того, какие там данные и как они обоснованы. Так что там в статье Ребекки Канн 1987 года? Что легло в основу новой религии? С чего все это началось? Давайте посмотрим. Основополагающая статья Канн и др. (1987) о «выходе из Африки» Во введении к статье – ни слова про Африку и якобы зарождение там человечества. То есть статья позиционирует себя как первая в этом отношении. Экспериментальная часть статьи – определение нуклеотидной последовательности мтДНК из 147 женщин из пяти основных регионов: Африка – 20 человек (двое рождены к югу от Сахары, остальные – чернокожие жители США, обычно метисы с примесью Y-ДНК мужчин-европеоидов, но у этих 18 человек «предполагается африканская мтДНК, на что также указывает характер мутаций фрагментов мтДНК»); Азия (Китай, Вьетнам, Лаос, Филиппины, Индонезия, Полинезия/Тонга) – 34 человека; Европеоиды (Европа, Северная Африка, Ближний Восток) – 46 человек; Австралийские аборигены – 21 человек; Новая Гвинея – 26 человек. Все мтДНК были расщеплены на фрагменты с помощью ферментов-рестриктаз, получены в сумме 467 независимых участков мтДНК, из которых 195 имели различия хотя бы у одного человека из всех 147. Другими словами, было идентифицировано 195 полиморфных участков мтДНК. В среднем анализ проводили на 9% от всей мтДНК. В общем, для того времени, 25 лет назад, это была довольно продвинутая в техническом отношении работа. Далее провели попарные сопоставления полученных фрагментов ДНК между всеми 147 участницами, и нашли, что эти попарные различия варьируются от нуля до 1.3 мутаций на каждые 100 нуклеотидов (0 до 1.3% различий), с общей усредненной величиной 0.32% различий. Но надо было показать, что эти различия больше всего у африканцев, поэтому все пять популяций подразделили на кластеры, исходя из групп попарных различий в каждой популяции. Получилось, что 46 европейских мтДНК расходятся на 36 кластеров, 34 азиатских мтДНК расходятся на 27 кластеров, 21 австралийских мтДНК расходятся на 15 кластеров, 26 мтДНК из Новой Гвинеи расходятся на 7 кластеров, а 20 африканских мтДНК определили в один кластер, постановив, что раз человечество вышло из Африки, то там должен быть всего один кластер. Так и записали в примечании к таблице в статье, где у всех указано много кластеров, а у африканцев всего один. Если кто понимает, что сделали авторы статьи, то желание читать дальше уже пропадает. Потому что дальше авторы стали определять, какая популяция старше других, для того на кластеры и разбивали. Но фокус был в том, что возраст каждой популяции расчитывали из разнообразия между кластерами внутри каждой популяции, а у африканцев он автоматически был задан самый большой, потому что у них кластер всего один. У всех разнообразие гасилось в каждом кластере, а у африканцев ничего не гасилось, кластер – один. Так и получилось, вот «разнообразия» по кластерам: Африка: 0.36% Азия: 0.21% Австралия: 0.17% Новая Гвинея : 0.11% Европа: 0.09% Далее авторы перевели эти «разнообразия» в хронологические показатели, а именно в года, когда эти территории были впервые заселены. Для этого взяли следующие цифры для калибровки: заселение Австралии произошло 40 тысяч лет назад, заселение Новой Гвинеи 30 тысяч лет назад, заселение Америки 12 тысяч лет назад, и получили, что мутации в мтДНК возникают со средней скоростью 2-4% (то есть 2-4 мутации на каждые 100 нуклеотидов) на миллион лет. Отсюда авторы статьи вычислили средний «возраст» кластеров в популяция: Африка: 90-180 тысяч лет Азия: 53-105 Австралия: 43-85 Новая Гвинея : 28-55 Европа: 23-45 Сделали топорно, но цифры получились довольно разумные (в пределах 100%-ной погрешности). Как показали последующие исследования уже других авторов, в том числе и мои, о которых будет рассказано ниже, африканские ДНК-линии пошли начиная примерно с 160 тысяч лет назад, плюс несколько архаичных африканских линий (гаплогруппы А0 и А00) возрастом примерно 180 и 210 тысяч лет, соответственно; азиатские и европейские линии – начиная с 64 тысяч лет назад, Австралия – примерно с 45-50 тысяч лет назад, а древнейшие костные остатки современного человека в Европе датируются 45 тысяч лет назад (Benazzi et al, 2011; Higham et al, 2011). Понятно, что авторы провели расчеты с точностью плюс-минус 100%, но тем не менее общая картина схвачена относительно верно. Подобным же образом авторы рассчитали, что общий предок всех мтДНК жил 143-285 тысяч лет назад, а поскольку общий предок всех африканских мтДНК жил, по их расчетам, 90-180 тысяч лет назад, то есть древнее всех (хотя и пересекается по возрасту в пределах погрешности расчетов), стало быть, тогда же он и вышел из Африки. Замечаете подмену понятий? Авторы расчитывают, что люди вне Африки произошли от более недавнего общего предка, и постулируют, что, значит, он вышел из Африки. В итоге, заключают авторы и это же пишут в Абстракте, женщина, общий предок всех мтДНК на планете, как «постулируется» (!), жила 200 тысяч лет назад (это уже трансформация величины 143-285 тысяч лет назад), и «вероятно» (!) она жила в Африке. Вот с этой статьи все и началось. Повторяю, что я ума не приложу, как такая статья смогла пройти рецензентов и быть опубликована в журнале Nature, с этими «постулируется» и «вероятно», и при отсутствии каких-либо данных о выходе современного человечества из Африки, но именно так эта статья и стала восприниматься и средствами информации, и в популяционной генетике, а оттуда и в науке и среди обывателя – что, значит, неопровержимо доказано, что современный человек вышел из Африки. Других генетических доказательств фактически не было, да и зачем? Все ведь уже доказано, не так ли? Принципиальные ошибки творцов и сторонников концепции «выход человечества из Африки» Есть основная ошибка, которая постоянно преследует популяционных генетиков. Если одна популяция генетически «разнообразнее» другой, то есть в совокупности древнее, они, как правило, считают, что она – предковая по отношению ко второй. Но это вовсе не так. Здесь надо смотреть совокупность факторов, а не перепрыгивать к выводу. Например, старший брат «разнообразнее» младшего, но это вовсе не значит, что младший – потомок старшего. Просто у них есть общий предок, их отец. Это же относится к разнообразным генеалогическим построениям, и если мы начнем сопоставлять племянников и отодвигать их общих предков к общему дедушке, прадедушке, пра-прадедушке и так далее, то увидим, что ветви потомков могут отходить от общего генеалогического дерева в разные времена, но нельзя их сравнивать «по возрасту» линейно, напрямую друг с другом, нужно непременно знать, когда жил их общий предок. Это ясно, если посмотреть на обычное дерево. Рядом сидят толстая ветка и молодая веточка, но вовсе не обязательно, что молодая выходит из старой. Зачастую они независимы до ствола, их общий предок – ствол, или ветка еще большей толщины. Концепция общего предка в статье 1987 года вообще не затрагивалась. Типичная ошибка попгенетиков – это «что вижу, то и пою». Раз сейчас живут в Африке – значит, всегда там жили. То, что общий предок африканцев и неафриканцев мог жить вне Африки, и в древние времена туда мигрировал – ими даже и не рассматривается. Есть еще одна принципиальная ошибка подхода попгенетиков, основанного на сравнении «разнообразия». Разнообразие информативно в данном смысле, как говорит термодинамика, только в замкнутых системах. Нью-Йорк значительно «разнообразнее», чем, скажем, Бостон, но означает ли это, что Бостон – потомок Нью-Йорка, вышел из него? Москва «разнообразнее» Новгорода, но потомок ли Новгород Москвы? Вовсе нет. Скорее, наоборот. Разнообразие получается зачастую от смешивания разных популяций, потому что система открытая. Вот в Нью-Йорке и в Москве намешалось, и накопилось большое «разнообразие». Африка – тоже открытая система. Туда в древности и в относительно недавнее время продвинулись многие миграции самых разных гаплогрупп, вот и «разнообразие». Даже гаплогруппа R1b в своей части продвинулась около 5 тысяч лет назад, теперь живут в Камеруне и Чаде (Cruciani et al, 2010), чернокожие, потому что смешались с местными красавицами. А гаплогруппа так и осталась, R1b. Добавили они в африканское «разнообразие»? Безусловно, как и множество подобных миграций в Африку. Время от времени появляются академические статьи, в которых описывается «вход в Африку». Последняя статья – в том же журнале Nature в августе 2013 года (Hayden, 2013), в которой описаны только что выявленные миграции популяций в Африку 3000 лет назад и 900-1800 лет назад. Добавили они к «разнообразию»? Безусловно. Тем более что ушли к югу от Сахары, откуда и брали пробы мтДНК авторы статьи 1987 года. Такую же ошибку делает Аткинсон в недавней статье (Atkinson, 2011), в которой он пишет: «генетическое и фенотипическое разнообразие уменьшается с удалением от Африки… что поддерживает гипотезу об африканском происхождении человека». Посмотрим на диаграмму, которая будет пояснена ниже. Слева – африканская ветвь, справа – неафриканская. Разнообразие (то есть древность) уменьшается слева направо, но вовсе не потому, что левая ветвь – предковая. Они обе выходят из общего предка, который, как будет показано ниже, в Африке не жил. Еще иллюстрация с падением разнообразия с удалением от Африки. Возраст гаплогрупп А в Африке – примерно 160 тысяч лет, после расхождения от альфа-гаплогруппы. Возраст гаплогрупп R1a и R1b на удалении от Африки – 20 тыс лет и 16 тыс лет, соответственно, они образовались в Центральной Азии (Klyosov and Rozhanskii, 2012a; Klyosov, 2012). Разнообразие падает от Африки к Центральной Азии? Падает. Потому ли, что R1a и R1b произошли от африканских гаплогрупп А? Совершенно нет. Это – несвязанные события и системы. Аналогия – если в одной части города находится дом престарелых, то «разнообразие» в нем самое высокое. Означает ли это, что все в городе, включая детский сад напротив, произошли от дома престарелых? Вовсе не обязательно. Это – несвязанные события и системы. Это могло бы быть верно, если бы система была замкнутой, то есть в город никто не въезжал на протяжении сотен лет. Но в реальности в город въезжают тысячи людей, которые не имеют никакого отношения к тем, кто в доме престарелых и кто в детском саду, да и в первый привозят престарелых со всей страны и из-за рубежа. Хотя если измерить – разнообразие в нем самое высокое. А вот предковости нет. Вот примеры «линейного мышления» в отношении «разнообразия»: …Разнообразие гаплотипов самое высокое в Африке (Hellenthal et al., 2008); Африка имеет не только самый высокий уровень генетической вариации в мире, но таже имеет значительное разнообразие в языках, культуре, природных условиях (Campbell and Tishkoff, 2010). Все это так, но к главному тезису о происхождении человечества якобы в Африке отношения не имеет. Это – всего пара примеров из сотен такого же рода. Для обсуждения «генетического разнообразия» надо знать генезис популяций, их историю, а не просто формально это «разнообразие» измерять у разных популяций и «линейно» сопоставлять. Это, повторяю, бич популяционных генетиков. Почему это? Слабая научная школа, другого объяснения нет. 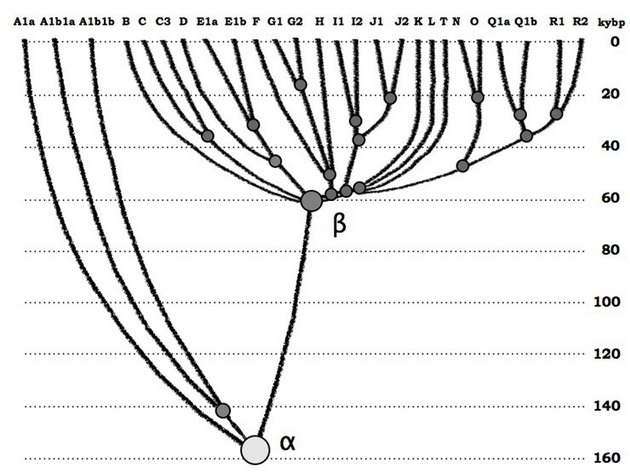 Диаграмма эволюции гаплогрупп современного человечества. На горизонтальной оси – основные гаплогруппы Y-хромосомы человечества, на вертикальной – абсолютная шкала времени. Общий предок альфа-гаплогруппы жил примерно 160 тысяч лет назад, общий предок бета-гаплогруппы (или гаплогрупп от В до Т) – 64±6 тысяч лет назад (из статьи A.A. Klyosov and I.L. Rozhanskii, Advances in Anthropology, 2012b). На диаграмме не показаны архаичные африканские линии А00 и А0 (последняя сейчас заменила в номенклатуре линии A1b на диаграмме слева), обновленное дерево гаплогрупп будет показано ниже. Есть еще (как минимум) одна причина более низкого «разнообразия» у неафриканцев. Примерно 64 тысячи лет назад их предки прошли «бутылочное горлышко популяции». Иначе говоря, в результате какого-то катаклизма неафриканцы почти все погибли, или выродились, и выжила их только небольшая группа. Образно говоря, выжили в конечном итоге потомки всего лишь одной пары, и вот к ним сходятся все генеалогические линии практически всех мужчин на планете. Что за катаклизм или другая напасть, как, например, мор, приключилась – неизвестно, и наибольший вес имеют две гипотезы – извержение вулкана Тоба, самое крупное из известных в истории человечества, примерно 70 тысяч лет назад, и похолодание в северной гемисфере. Климатологи утверждают, что ранг катастрофичности похолодания выше, чем ранг извержения Тобы. Как там ни было, вот что получилось (см. приведённую диаграмму). Понятно, что если измерять «разнообразие» ветвей слева (африканской по нынешнему проживанию ее представителей) и справа (неафриканской, тоже по нынешнему проживанию), то первая будет древнее. Но правая ветвь не выходит из левой, у них – общий предок, альфа-гаплогруппа. Как видно, диаграмма объясняет все результаты статьи Канн 1987 года, но без выхода из Африки. Другие доказательства правильности этой диаграммы будут приведены ниже. Умножение фантазий в ходе распространения концепции «выхода человечества из Африки» Со временем как-то из воздуха появилась цифра, что современный человек вышел из Африки 70 тысяч лет назад, и ее тоже стали цитировать в сотнях академических статей, так что и концы затерялись, кто и когда это сказал первым. Да и кого это интересовало? Вышел современный человек из Африки? Вышел, давно и неопровержимо показано, еще в 1987 году. Когда вышел? Так вот же, все пишут, что 70 тысяч лет назад, тоже давно и неопровержимо показано. Какие еще вопросы? Кто там в консенсус не верит? Давайте-ка на такого посмотрим, а то и меры примем. И пошли десятки и сотни академических статей, первая фраза в которых обычно была – «Как известно, анатомически современный человек вышел из Африки примерно 70 тысяч лет назад». Впрочем, и эта датировка была «плавающей», и ниже даются примеры разных датировок «выхода из Африки» в разных статьях. Небольшой секрет – ни одна из них не была действительно расчетной. Все они – просто так, «с потолка». Да и не было до последнего времени соответствующего расчетного аппарата, а тот, что был – читатель уже увидел, какой он и какая там точность. 50 тысяч лет назад (Jobling & Tyler-Smith, 2003); 50 тысяч лет назад (Thomson et al, 2000); 50-60 тысяч лет назад (Shi et al., 2010); 50-60 тысяч лет назад (Mellars, 2011); 50-70 тысяч лет назад (Hudjasov et al., 2007); 50-70 тысяч лет назад (Stoneking & Delfin, 2010); 60 тысяч лет назад (Li & Durbin, 2011); 60 тысяч лет назад (Henn et al., 2011); 60 тысяч лет назад (Wei et al., 2013); 60-70 тысяч лет назад (Ottoni et al., 2010); 60-80 тысяч лет назад (Forster, 2004); 54±8 тысяч лет назад (Forster et al., 2001); 60 тысяч лет назад (Stewart & Stringer, 2012); 45-50 тысяч лет назад (Fernandes et al., 2012); 50-65 тысяч лет назад (Behar et al., 2008); 50-60 тысяч лет назад (Cann, 2013); 60 тысяч лет назад (Chiaroni et al., 2009); 50-75 тысяч лет назад (Patin et al., 2009); 50 тысяч лет назад (Edmonds et al., 2004); 45 тысяч лет назад (Moorjani et al., 2011); 50-70 тысяч лет назад (Xue et al., 2005); 70-80 тысяч лет назад (Majumder, 2010); 40 тысяч лет назад (Campbell and Tishkoff, 2010); 50 тысяч лет назад (Poznik et al, 2013); 55-70 тысяч лет назад (Soares et al., 2009); между 40 и 70 тысяч лет назад (Sahoo et al., 2006); между 35 и 89 тысяч лет назад (Underhill et al., 2000); между 80 и 50 тысяч лет назад (Yotova et al., 2011); между 50 и 100 тысяч лет назад (Hublin, 2011); между 27-53 и 58-112 тысяч лет назад (Carrigan and Hammer, 2006); 70-60 тысяч лет назад (Curnoe et al., 2012); ∼ 110 тысяч лет назад (Francalacci et al, 2013); 200 тысяч лет назад (Hayden, 2013). На самом деле никакой даты обоснованно привести нельзя. Ее просто нет. Да и все эти датировки выше никому не нужны, они ничего не дают и не отвечают по сути ни на какой вопрос. Это – все та же мантра. Что говорят на самом деле экспериментальные данные и их более широкая интерпретация? Остановимся на время с критикой, и посмотрим – а что же есть? Если современные неафриканцы – не потомки древних африканцев, то откуда это следует? Чьи они потомки? Археологические, палеонтологические данные по Африке придется с сожалением отбросить. Они познавательны из общих соображений, но мы не знаем, были ли у найденных костных остатков вообще выжившие потомки. Возможно, мы наблюдаем остатки терминировавшихся линий. Пока не будут установлены гаплогруппы и гаплотипы тех костных остатков, они нам ничего не скажут о преемственности этих эволюционных линий. Далее, мы не знаем, откуда эти костные остатки там появились. Возможно, их близкие предки мигрировали в Африку. Действительно, если возможно было выйти из Африки, то так же возможно было туда и войти. Более того, известны многие примеры миграций в Африку. Многие датировки древних костных остатков неверны, и примеры будут даны ниже. Многие заявленные как «древние Homo sapiens» имеют выраженные архаичные признаки, и их отнесение к Homo sapiens вообще является спорным или просто неверным. Многие находки относятся вообще не к костным остаткам, а к стоянкам, пещерам, найденным там раковинам, каменным орудиям труда. Неизвестно, кто там вообще был, и найденная там охра тоже ни о чем не говорит. Неандертальцы Евразии тоже применяли каменные орудия и охру для своих целей. Так что к рассматриваемому вопросу о том, как соотносятся древние африканцы и неафриканцы, больше подходит расмотрение ДНК тех и других. Если эти данные поддерживаются и археологией-антропологией – это замечательно, но пока таких данных мало, если они вообще есть. Посмотрим на них. Данные по ДНК можно рассматривать в трех вариантах, которые в принципе обязаны давать взаимно согласованные данные. Это (1) гаплотипы и гаплогруппы Y-хромосомы человека, (2) мтДНК человека, и (3) геном человека. Последнее фактически означает картину необратимых мутаций в ДНК, которая поддается интерпретации в отношении эволюции человека, показывая направление перетекания мутаций и появления новых в ходе эволюционного развития. Например, в геноме как неандертальца, так и современного человека есть много одних и тех же мутаций, которые есть и в ДНК шимпанзе. Это означает, что данные мутации – от общего предка человека и шимпанзе. Но если в нас есть и мутации от неандертальца, которых нет у шимпанзе – то это может означать, что неандерталец – наш прямой предок. Такие мутации или не выявлены, или их очень немного и они спорные. Данные сейчас пересматриваются. Недавно провозглашенные 1-4% как якобы у современного человека от неандертальца тоже сейчас пересматриваются. Скорее всего, они неверные. Таким же образом как в неафриканцах, так и в африканцах есть общие мутации от общего предка с шимпанзе. Их много, и они неинтересны при решении вопроса о том, произошли ли мы от африканцев. Эти мутации должны быть отфильтрованы. А вот есть ли в нас такие мутации, которые есть у африканцев, но нет у шимпанзе – вот это вопрос, на который должен быть ответ. Этот ответ, если такой будет получен, должен согласовываться с данными по гаплотипам и гапллогруппам Y-хромосомы и по мтДНК. Вот так должно строиться исследование того вопроса, каковы эволюционные пути современного человечества. На самом деле, этот ответ уже получен – нет в нас «африканских» мутаций, приобретенных ими за последние 150-200 тысяч лет. Мутации от общего предка с шимпанзе, которым миллионы лет, в наших ДНК есть, и много, а мутаций от африканцев, приобретенных ими за последние 160 тысяч лет, в наших ДНК нет. Об этом и пойдет речь. Итак, что говорят на этот счет экспериментальные данные? Начнем с гаплотипов и гаплогрупп человека, проецируемых на времена более 100 тысяч лет назад. Проецируемых – потому что ископаемых гаплотипов и гаплогрупп того времени нет. Пока не выявлены, задача технически очень сложная, потому что за такие времена ДНК человека разлагается почти полностью, особенно под влиянием микроорганизмов. ДНК неандертальца (точнее, неандерталки) давностью 45 тысяч лет выделена и в значительной степени расшифрована, а вот ДНК человека 160 тысяч лет назад – это задача на порядки потруднее. Как поступают в таких случаях? Определяют гаплотипы у современных популяций мужчин, если анализ проводят по Y-хромосоме. Но выборку анализируют не по какой-то «новогвинейской» или «африканской» популяции, которая может быть совершенно разнородной, а среди носителей определенного субклада гаплогруппы, то есть среди людей, объединенных конкретным набором мутаций. Они – родственники, и для них довольно точно рассчитывается, когда жил их общий предок. Например, среди современных африканцев есть довольно представительная группа людей, которая по классификации попадает в субклад A1b1b2b, помеченный цветом (третий снизу) на дереве субкладов гаплогруппы А ниже. Это дерево показывает иерархию субкладов гаплогруппы A, то есть эволюцию гаплогруппы А. Видно, как ветвится дерево – от ствола в сторону отошла древнейшая гаплогруппа А00, ее ветви (субклады) пока неизвестны. Ствол продолжает гаплогруппа А0-Т, которая разошлась на два субклада – А0 и А1; А1 в свою очередь разошлась на А1а и A1b; A1b – на A1b1 и ВТ. Сводная гаплогруппа ВТ, как будет ещё показазано, очень удалена от гаплогрупп серии «А», да и в серии «А» непонятно, какие субклады могут быть названы африканскими по происхождению. Пока складывается, что только гаплогруппы А00 и А0, то есть первая и третья сверху (отошедшие в сторону от стволовой ветви, ведущей от общего предка с шимпанзе миллионы лет назад, к неафриканской гаплогруппе ВТ) на показанном ниже дереве, и субклады последней (A0a, A0b, A0a1, A0a2, A0a1a и A01ab) могут рассматриваться как африканские по происхождению или по прибытию в Африку более 100 тысяч лет назад. Остальные, начиная с А1 (находящейся на той же стволовой ветви), раздваиваются на африканские (отходящие в сторону) и предположительно неафриканские (стволовые) ветви.  Давайте рассмотрим еще раз, более четко, как ветвится дерево гаплогрупп, как каждая ветвь расходится на вилки, и как одна часть вилки уходит (мигрирует) в Африку, а другая часть остается вне Африки, и опять расходится на очередную вилку. Иначе говоря, в Африку миграции приходили волнами. В итоге прослеживается неафриканский ствол, ведущий к нам с вами, читатель, и от которого в сторону отходят африканские побеги. Мы от этих побегов не произошли. Здесь надо заметить, что термины «ствол», «стволовая» и «отходящие в сторону» выбраны условно, и можно, наоборот, назвать африканские ветви стволовыми, а неафриканские – отходящими в сторону. Эти понятия на самом деле симметричны. Вилка 1 – от основного эволюционного Y-хромосомного «ствола», идущего от общих предков с приматами (шимпанзе, горилла, орангутанг, макака) примерно 300-600 тысяч лет назад отходит ветвь неандертальцев (Homo neanderthalensis); они не были африканцами, во всяком случае, их следов в Африке не обнаружено, поэтому можно полагать, что и общий ствол 300-600 тысяч лет назад был неафриканским родом Homo. Вилка 2 – от ствола примерно 210 тысяч лет назад отходит наиболее древняя из пока обнаруженных ветвей букета гаплогрупп А, гаплогруппа A00 (сейчас все ее обнаруженные носители живут в Африке в составе племени Mbo, или являются афро-американцами; никаких сведений об их антропологии или анатомии обнаружить не удалось, в статье, приводящей их гаплотипы, об этом ни слова). Вилка 3 – ствол достигает гаплогруппы А0-Т (предположительно неафриканской), которая расходится примерно 180 тысяч лет назад на африканскую гаплогруппу А0 и предположительно неафриканскую А1; другими словами, от ствола отходит очередная африканская гаплогруппа А0. Вилка 4 – Неафриканская гаплогруппа А1 расходится на африканскую А1а и предположительно неафриканскую А1b; другими словами, от ствола отходит очередная африканская гаплогруппа А1а. Вилка 5 – Неафриканская гаплогруппа A1b расходится на африканскую А1b1 и неафриканскую BT (бета-гаплогруппа на первой диаграмме); другими словами, от ствола отходит очередная африканская гаплогруппа А1b1. Теперь – очень важное положение нашего рассмотрения. Вилки 3, 4 и 5 расходятся от гаплогрупп А0-Т, А1 и А1b, соответственно. От первой в сторону отходит А0 (которая найдена в Африке) и А1 (носители которой пока нигде не найдены). Мы, неафриканцы, происходим от А1 (и не происходим от А0, ее мутаций в нас нет). От А1 в сторону отходит А1а (которая найдена в Африке) и A1b (носители которой пока нигде не найдены). Мы, неафриканцы, происходим от нее. В нашей Y-хромосоме – мутации от A1b, но не от A1a. От A1b в сторону отходит A1b1 (которая найдена в Африке, Европе и Азии) и BT, из которой вышли все неафриканские гаплогруппы, в том числе основные европейские гаплогруппы R1a, R1b, I1, I2, N1c1. Для того, чтобы «доказать», что все люди на Земле вышли из Африки (в виде своих предков, естественно), сторонники концепции «выхода из Африки» объявляют все эти три узловые гаплогруппы – A0-T, A1 и A1b «африканскими». Повторяю, что никакая из них в Африке не найдена. Но это «сторонников» не беспокоит. Читатель уже понял, что там – другие приемы, которые научными назвать никак нельзя. Они объявляются африканскими, и «сторонники» говорят – ну, смотрите, все европейские и азиатские гаплогруппы выходят из африканских, из А0-T, A1 и A1b. Всё, концепция «выхода из Африки» доказана. На самом деле, это не доказательство, а насмешка над научным рассмотрением и над здравым смыслом. Значительно более вероятно, что эти три гаплогруппы вовсе не африканские, а их носители жили вне Африки. Тогда легко объясняется связка между предком светлокожего неандертальца (об этом – ниже) и светлокожими же современными людьми. Легко объясняются уходы в Африку – после расхождений-вилок – носителей гаплогрупп А0, А1а, A1b1, которые и сейчас преимущественно живут в Африке. Легко объясняются огромные временные дистанции между африканскими и неафриканскими гаплогруппами, потому что они сходятся к далеким общим предкам, а не выходят непосредственно друг из друга (тогда дистанции были бы примерно 60-70 тысяч лет, а они на самом деле 250-300 тысяч лет. Принципиально не могут выйти неафриканские линии из африканских, чтобы их разделяли 250-300 тысяч лет. Да и сами сторонники «выхода из Африки» постоянно провозглашают, что выход был 60-70 тысяч лет назад. Они же не считали, и не знают, что там дистанция на самом деле в 4-5 раз больше. Поэтому в описании вилок выше я везде пишу «предположительно неафриканская гаплогруппа» A0-T, A1, A1b. Таким образом, где бы не жили первопредки неандертальцев и те, с кем они разошлись в ходе эволюции (то есть те, которые продолжали «основной ствол» эволюционного дерева Y-хромосомы), носители гаплогрупп А00, А0, A1a, A1b1 мигрировали от них в Африку, и продолжали свою эволюцию там, принимая многочисленных более поздних мигрантов в Африку и тем самым увеличивая африканское «разнообразие». В целом, можно насчитать четыре основных древних миграции в Африку на протяжении последних нескольких сот тысяч лет – гаплогруппа А00 примерно 210 тысяч лет назад, гаплогруппа А0 примерно 180 тысяч лет назад, гаплогруппа А1а примерно 160 тысяч лет назад, гаплогруппа A1b1 примерно 70 тысяч лет назад. Разумеется, происходили и более поздние миграции, например 3000 и 900-1800 лет назад, описанные в работе (Hayden, 2013), которые тоже увеличивали «генетическое разнообразие» в Африке, так что «разнообразие» – не аргумент «прародины». Я уже упомянул, что носители гаплогруппы A1b1 живут и в Африке, и в Европе, и в Азии. Видимо, поэтому субклад A1b1b2b-M13 в Проекте гаплогруппы А оказывается наиболее многочисленным. Он расходится на две основные ветви – арабскую и европейскую. Кто был родоначальником этих ветвей и где он жил – мы не знаем, но ветвь довольно неглубокая, то есть она относительно недавно прошла бутылочное горлышко популяции. Ее гаплотипы представляют собой ценный источник информации, поскольку помещают даже неглубокую (по времени) ветвь в поле всех гаплотипов человечества. Гаплотипы и после бутылочного горлышка популяции не могли самозародиться, они могли только продолжить эволюцию от древнейших общих предков. Анализ гаплотипов с использованием самых «медленных», самых стабильных 22 маркеров Y-хромосомы (Klyosov, 2011), показывает, что общий предок арабской ветви субклада A1b1b2b имел гаплотип 12 11 11 9 11 10 10 9 12 12 7 12 8 0 13 11 16 9 14 9 11 11 а общий предок европейской ветви имел гаплотип 12 11 11 9 11 10 10 9 12 12 7 10 8 0 13 11 16 10 14 9 11 11 Между ними всего три мутации, что помещает общего предка арабской и европейской ветвей примерно на 7170 лет назад, с погрешностью плюс-минус пять процентов. Для целей нашего описания эти расчеты пока не очень важны, поскольку видно, что приведенные выше гаплотипы близки друг к другу. Давайте сопоставим эти гаплотипы с предковым африканским гаплотипом группы A00: 13 11 12 10 11 16 10 9 14 14 8 8 8 9 12 11 12 8 12 12 11 11 Это сопоставление показывает разницу уже в 30 и 29 мутаций, то есть разводит общих предков этих гаплотипов как минимум на 286-308 тысяч лет (формулы расчетов опубликованы в работе [Klyosov, 2011, 2012]), и помещает общего предка гаплогруппы А00 примерно на 210 тысяч лет назад. Зависимость между числом мутаций и временем не линейная, а степенная, поскольку на больших временах часть мутаций возвращаются обратно, и на это при расчетах вводится соответствующая статистическая поправка (Klyosov, 2009; Klyosov, 2012). Гаплотипы гаплогруппы А00 получены у чернокожего племени Мбо, живущего в Камеруне, и у афро-американца, предположительно вывезенного столетия назад из того же племени (Mendez et al, 2013). Если мы теперь сравним эти гаплотипы с предковым гаплотипом гаплогруппы В 11 12 11 11 11 10 11 8 16 16 8 10 8 12 10 11 15 8 12 11 12 11 то мы увидим 29 мутаций от гаплогруппы А00, и практически столько же – 29 и 27 мутаций – от арабской и европейской ветвей гаплогруппы A1b1b2b. Это – как минимум 286-248 лет между общими предками гаплогрупп А и В. Это колоссальное разделение во времени никак не позволяет гаплогруппе В быть потомком гаплогруппы А. А вот иметь общего предка 160 тысяч лет назад и разойтись от него на 250-300 тысяч лет – можно. Это опять согласуется с диаграммой выше. Нельзя эти гаплогруппы сопоставлять «линейно» только потому, что они находятся визуально рядом на дереве гаплотипов, как нельзя сопоставлять ветви дерева в лесу «линейно», по расстоянию между ними, только потому, что они оказались рядом. А рядом могут оказаться ветви березы и ели, растущей по соседству. Итак, гаплогруппа В очень удалена от гаплогруппы А, на 27-29-30 мутаций. А вот от европейских (в значительной степени) гаплогрупп R1a и R1b она удалена не так сильно, всего на 12 и 10 мутаций, соответственно: 11 12 13 11 11 12 11 9 15 16 8 10 8 12 10 12 12 8 12 11 11 12 (R1b-M269) 12 12 11 11 11 11 11 8 17 17 8 10 8 12 10 12 12 8 12 11 11 12 (R1a-Z280) Сами же эти гаплотипы (R1b и R1a) разделены всего 8 мутациями, что соответствует времени жизни их общего предка (гаплогруппа R1) примерно 26 тысяч лет назад. Общий предок гаплогруппы В жил около 50 тысяч лет назад, и он не образовался из гаплогруппы А, они – независимые ДНК-генеалогические линии, выходящие из одного общего предка – альфа-гаплогруппы, 160 тысяч лет назад. В Европе живут и другие носители гаплогруппы А, правда, их пока найдено немного. Несколько лет назад вышла научная статья под названием «Африканцы в Йоркшире?» (King et al, 2007), в которой описано семейство носителей гаплогруппы А в Англии, которые и не подозревали, что у них африканские корни по мужской линии. Базовый 17-маркерный гаплотип у них оказался следующий (в порядке маркеров DYS393, 390, 19, 391, 388, 439, 389-1, 392, 389-2, 437, 438, 434, 435, 436, 460, 461, 462): 14 23 17 10 10 11 12 11 17 14 8 12 12 11 11 12 12 А у арабской ветви субклада A1b1b2b, описанного выше 13 21 15 9 11 12 13 11 18 16 10 9 11 11 11 13 13 Между ними – 20 мутаций на 17 маркерах, что соответствует как минимум 19 тысячам лет до их общего предка, английских и арабских гаплотипов гаплогруппы А. Кто в этом случае куда передвигался – или в Африку, или из Африки – сказать невозможно. Сценарии могли быть любые. Сторонник же концепции «выход из Африки» тут же скажет, что они вышли из Африки. Вот такая психологическая установка. Дебаты последних двух лет в отношении приведённой диаграммы Когда статья, содержащая представленную выше диаграмму и её интепретацию, была опубликована в мае 2012 года в журнале «Advances in Anthropology» (Успехи антропологии), это поначалу вызвало неприятие у популяционных генетиков. Конкретно, неприятие вызвали три основных вывода: (1) африканские и неафриканские ДНК-линии разошлись примерно 160 тысяч лет назад, и между ними имеется соответствующее значительное расстояние; (2) неафриканские ДНК-линии не являются потомками африканских гаплогрупп А00, А0, А с субкладами; и, как следствие, (3) никакого «выхода из Африки» у современного человечества не было, во всяком случае в последние 200 тысяч лет. А если и было, то было туда-сюда, встречными миграциями, и современного человечества очередная «туда-сюда» миграция не породила. Во всяком случае, в данном отношении они эквивалентны. Надо сказать, что это неприятие в академической научной печати вообще не высказывалось. Популяционные генетики как воды в рот набрали. Страсти кипели на англоязычных форумах, в неформальных дискуссиях. Было объявлено, что эта диаграмма и, соответственно, ее выводы полностью противоречат консенсусу про выход человечества из Африки, и противоречат всем опубликованным диаграммам и деревьям гаплогрупп, полученным с помощью геномных исследований. Было также объявлено, что неафриканские линии выходят из гаплогрупп с индексом «А», значит, африканских. Было объявлено, что это не согласуется с эволюцией женских, мтДНК, у которых (ныне) неафриканские линии тоже выходят из Африки, а мужские и женские гаплогруппы должны были выходить из Африки вместе. На самом деле, эти все возражения были в принципе неверными. Оппоненты или не хотели, или не могли разобраться, и, как обычно, пытались «брать горлом». Давайте посмотрим и убедимся, что никаких противоречий на самом деле нет, и оппоненты просто повторяли заученные мантры, что в популяционной генетике слишком часто принято.
|
|
#4
22.03.2014, 11:41
|
||||
|
||||
|
Недавняя книга «Эволюционная генетика человека» –
правильные данные, неверная интерпретация Раскрываем новую книгу – «Эволюционная генетика человека» (Human Evolutionary Genetics), авторы Jobling, Hollox, Hurles, Kivisild, Tyler-Smith, год издания 2014 (именно так, издательство забежало на полгода вперед), глава 9 – «Происхождение современного человека», стр. 304-305. Раздел «Митохондриальные ДНК». Цитата: «Исследования показали поразительные особенности: полное разделение африканских и неафриканских ДНК-линий» («Complete separation of African and non-African lineages»). Раздел «Y-хромосомы». Цитата: «Хотя менее детальные, чем мтДНК, исследования показали близкие параллели: полное разделение африканских и неафриканских ДНК-линий» («Complete separation of African and non-African lineages»). Как мы видим, никаких противоречий с диаграммой выше. Но авторы уже накручивают свои интерпретации, основываясь на данных 2000 года – и по мтДНК, и по Y-хромосоме. Так, Y-хромосомную гаплогруппу В считают африканской, и пишут, что соответствующая ветвь содержит «как африканские, так и неафриканские ДНК-линии». Смотрим на диаграмму – да, гаплогруппа В находится в одном кусте с неафриканскими гаплогруппами, и мы выше показали, что она удалена от африканских гаплогрупп, и находится в одном кластере с неафриканскими, с одним общим предком. А почему авторы назвали ее «африканской»? Да так, многие носители гаплогруппы В сейчас живут в Африке. Помните, как я писал про популяционных генетиков? «Что вижу, то и пою». У них раз в одном кластере находятся как гаплогруппа В, так и неафриканские линии, значит, «выход из Африки». А они все там, в том кластере, неафриканские. Да если бы и была неафриканская линия вместе с неафриканскими – почему непременно «выход из Африки»? Почему с тем же успехом не «вход в Африку»? А так, маршируют по накатанному тракту, ответ заранее известен. Датировку этого кластера, содержащего неафриканские гаплогруппы и гаплогруппу В (тоже неафриканскую по происхождению) авторы книги дают как 52±28 тысяч лет назад. У меня в статье – 64±6 тысяч лет назад. Где же противоречие? Те же авторы приводят датировку всех ДНК-линий – 172±50 тысяч лет назад. Действительно, у меня в статье 160±12 тысяч лет назад. Где же противоречие? То есть популяционные генетики оспаривают не по сути, не с данными в руках, а просто «в принципе», ради неприятия. Обычное дело. В отношении мтДНК авторы симметрично приводят те же самые интерпретации, что и с Y-хромосомой – похожая ветвь, содержащая «африканские мтДНК» (потому что сейчас там живут) и неафриканские – значит, «выход из Африки», причем датировка этой «смешанной» ветви между 31 и 79 тысяч лет назад, с медианой 40 тысяч лет назад, датировка совокупности всех мтДНК – между 40 и 140 тысяч лет назад, медиана 59 тысяч лет назад. Разнобой в датировках между Y-хромосомными данными и мтДНК авторы и не обсуждают, а зачем? Вывод давно готов – «выход человечества из Африки». Этот же вывод, хотя в осторожном виде – и в заключении по главе. Там же и про «более высокое генетическое разнообразие в Африке», и то, что в Африке человек появился примерно 200 тысяч лет назад, а вне Африки – после 45 тысяч лет назад. Здесь же и про консенсус специалистов о «выходе из Африки». Мы видели, что все эти (или подобные) датировки и «разнообразия» объясняются диаграммой выше, но попгенетики других объяснений не хотят. У них – «консенсус». История с продолжением статьи Канн (1987) «о выходе из Африки», но уже без Канн (1991) По-своему интересно продолжение статьи Канн и других (Cann, Stoneking and Wilson, 1987), которую мы обсуждали выше. Новая статья вышла спустя четыре года (Vigilant et al, 1991), в авторах Канн уже нет, но есть два прежних соавтора, Стоункинг и Уилсон, с тремя новыми авторами. Статья 1991 года сообщает, что работа Канн и др. (1987) встретила резкое неприятие многих специалистов из-за того, что общий предок человечества якобы жил в Африке, и признает, что в статье Канн и др. (1987) было много слабых звеньев. Эти слабые звенья авторы (двое из которых и были авторами той слабой, по их признанию, работы) перечисляют на протяжении целого абзаца – там и непрямой метод сопоставления мтДНК, и малая выборка, причем состоящая в основном из американцев африканского происхождения, и заведомо непригодный метод «средней точки», примененный авторами статьи 1987 года, и отсутствие статистической обработки полученных данных, и «неадекватная калибровка» скорости мутаций в мтДНК, и другие. Иначе говоря, эта заведомо слабая статья, по признанию самих авторов, легла в основу теории «выхода из Африки». Но процесс уже пошел, поэтому последующая статья (1991) преследовала цель все-таки оправдать концепцию «выхода из Африки», что неафриканцы произошли от африканцев, и фактически заменить собой слабую, раскритикованную статью 1987 года. И в чем было то самое оправдание? Показать, что африканские мтДНК древнее неафриканских. Но это опять продолжение той вечной фундаментальной ошибки популяционных генетиков, что если одна популяция древнее другой – то первая популяция якобы предковая по отношению ко второй. Взглянем опять на ту же самую диаграмму выше – левая ветвь древнее правой, но она не предковая по отношению к правой. У них – один общий предок. И вот эту фундаментальную ошибку попгенетики повторяют все последующие 25 лет, до настоящего времени. Опять и опять авторы статьи (1991) повторяют, что африканская ветвь древнее неафриканской, значит, предковая, не понимая, что это вовсе не является доказательством «предковости». Мой дядя «древнее» чем я, но он не мой предок. В заключении статьи (1991) авторы пишут: мы представили самые строгие доказательства, что наш общий предок жил в Африке 200 тысяч лет назад. На самом деле, как читатель уже давно понял, эти доказательства на самом деле были о том, что сохранившаяся ветвь людей, ныне живущих в Африке, древнее сохранившейся ветви людей, живущих вне Африки. О «предковости» эти «доказательства» ничего не говорят. Для этого надо сравнивать гаплотипы популяций (что авторы статьи 1991 года не делали, и попгенетики не делают до сих пор) и их снип-мутации (что авторы не делали), и последнее тоже показывает, что наши предки из Африки не вышли. Об этом – следующий раздел. Снип (SNP)-мутации показывают, что мы – не потомки африканцев гаплогрупп А или В Перейдем к недавней статье (Scozzari et al, 2012), которую часто ставят в пример как образцовую работу по геному африканцев и обоснованию «выхода человечества из Африки». Действительно, статья объявляет об обнаружении 22 новых необратимых мутаций в Y-хромосоме человека, о подтверждении 146 известных мутаций и о конструировании нового, улучшенного дерева гаплогрупп и субкладов африканцев с переходом в неафриканскую часть дерева, и именно сводной гаплогруппы СТ. Это – вся правая часть дерева выше на диаграмме, от гаплогруппы С до R2. Авторы статьи называют ее «вышедшей из Африки». Посмотрим, так ли это. Дерево гаплогрупп и субкладов из статьи (Scozzari et al, 2012):  Дерево самых древних гаплогрупп и субкладов, приведенное в статье (Scozzari et al, 2012). Показаны номера необратимых мутаций (SNP, Single Nucleotide Polymorphism, или снипы), которые задают определенные субклады. Видно, что больше половины субкладов относятся к гаплогруппе А, которую авторы считают африканской. Все остальные субклады, кроме одного, относятся к гаплогруппе В, которую авторы также считают африканской. Гаплогруппа в нижней правой части, СТ, состоит, по данным авторов, из 19 ДНК-линий, все неафриканские. Картинка кликабельна. Обратим внимание на некоторые особенности дерева на этом рисунке. Оно начинается (точнее, продолжает эволюционный ствол Y-хромосомы человека) в верхней левой части диаграммы, тут же идет первое расхождение, или вилка (гаплогруппа A0-T, хотя это название на диаграмме не показано), на гаплогруппу A1b (так на диаграмме) c субкладами, с одной стороны, и на остальную часть дерева, с другой. Иначе говоря, от дерева отходит первая африканская ветвь, и никакие неафриканцы (гаплогруппа СТ) от нее уже не происходят. В статье используется уже устаревшая номенклатура 2011 года, и то, что в статье обозначено как A1b, сейчас называется А0, со снипами V148, V149 и другими, приведенными на верхней линии диаграммы (см. выше также дерево гаплогруппы А). На следующей вилке (гаплогруппа А1) в сторону отходят африканские гаплогруппы А1а и А1а1 с их снипами М31, Р82, V4 и другими, в другую сторону – остальная часть дерева. От второй африканской ветви (A1a c cубкладом) неафриканцы (гаплогруппа СТ) тоже не происходят. Третья вилка – гаплогруппа А1b согласно нынешней классификации. От нее отходят в сторону африканские гаплогруппы А2 и А3 с субкладами (номенклатура устаревшая), сейчас это A1b1 cо снипом V249/L419, которая далее расходится на субклады A1b1a-V50 (бывшая А2) и A1b1b-M32 (бывшая А3), оба с подгруппами. Среди последних – субклад A1b1b2b-M13, тот самый, из которого выходят арабская и европейская ДНК-линии, гаплотипы которых мы рассматривали выше. Другая ветвь этой вилки от гаплогруппы А1b – сводная гаплогруппа ВТ, показанная в нижней части рисунка из статьи (Scozzari et al, 2012). Совершенно ясно, что эта гаплогруппа ВТ никоим образом не образуется от «африканских» гаплогрупп с индексом А, которые все находятся в верхней части этого рисунка. Слово «африканские» здесь приходится брать в кавычки, поскольку среди их субкладов и те самые европейская и арабская ветви, причем европейская – это, в основном, Англия, Ирландия, Шотландия, Турция (хотя лишь 3% Турции территориально находится в Европе), арабская – в основном Саудовская Аравия, и гаплотипы из Англии, Швейцарии, Финляндии и других стран. Естественно, можно утверждать, что европейские и азиатские гаплотипы гаплогруппы А когда-то вышли из Африки вместе с мигрантами оттуда, но с таким же успехом можно утверждать, что они попали в Африку тем же путем. Так что эти аргументы не проходят, хотя только «из Африки» вызывает поддержку среди сторонников «выхода из Африки». Альтернативные объяснения они не рассматривают в принципе. Завершая рассмотрение рисунка из статьи (Scozzari et al, 2012), стоит отметить и тот факт, что общепризнанно «неафриканская» сводная гаплогруппа СТ (нижняя линия на рисунке) не выходит и из гаплогруппы В с ее субкладами, назвать ее африканской или нет. Видно, что эволюционный путь гаплогруппы СТ минует все «африканские» гаплогруппы, даже если они все содержат европейских или других предков. Если двигаться вверх по временной шкале (то есть слева направо) мутационный путь гаплогруппы СТ выходит из основного «ствола» эволюции Y-хромосомы человека, то есть ниже «хвостика», обозначенного на рисунке слева вверху, проходит через гаплогруппу А0-Т (никаких сведений, что она якобы «африканская» – нет), далее через гаплогруппу А1 (то же самое, нет никаких сведений, что она якобы «африканская»), далее через гаплогруппу A1b, тоже неафриканскую, далее через гаплогруппу ВТ, и становится гаплогруппой СТ. Все три «африканские» ветви (А0, А1а, А1b1) на этом пути уходят в сторону путем соответствующих ответвлений, вилок. Мы остановились на этом столь подробно, потому что в академической литературе, и тем более в популярной, такой или подобный анализ никогда не проводился. Обычно показывается дерево, как на рисунке из статьи (Scozzari et al, 2012), и скороговоркой сообщается, что оно «свидетельствует о выходе человечества из Африки». Никакого объяснения не дается. Ответ готов уже заранее. Иногда доходит до абсурда – сообщается, что гаплогруппы ВТ и СТ выходят из гаплогрупп А1b, или A1, или А0-Т, значит, это африканские гаплогруппы, поскольку имеют индекс «А». То есть подмена понятий зашла настолько далеко, что абсолютно условные названия принимаются за доказательства тезиса. То, что эти гаплогруппы могли с таким же успехом назвать X, Y, Z, или W, в голову уже не приходит. Раз «А», значит, Африка, нет и сомнений. Так что никакого противоречия между рисунком из статьи (Scozzari et al, 2012) и диаграммой выше нет, они показывают те же самые закономерности эволюционного развития гаплогрупп, а именно расхождение на «африканские» и «неафриканские» ветви. Разница между ними только в том что на рисунке из статьи (Scozzari et al, 2012) более детально показаны субклады гаплогрупп А («африканской») и В, а на диаграмме – «неафриканские» гаплогруппы ВТ. Ещё в том, что первое изображение построено с учетом хронологической шкалы, а второе – без учета. Чтобы показать сходство, расположим оба дерева гаплогрупп вертикально, в одном направлении. 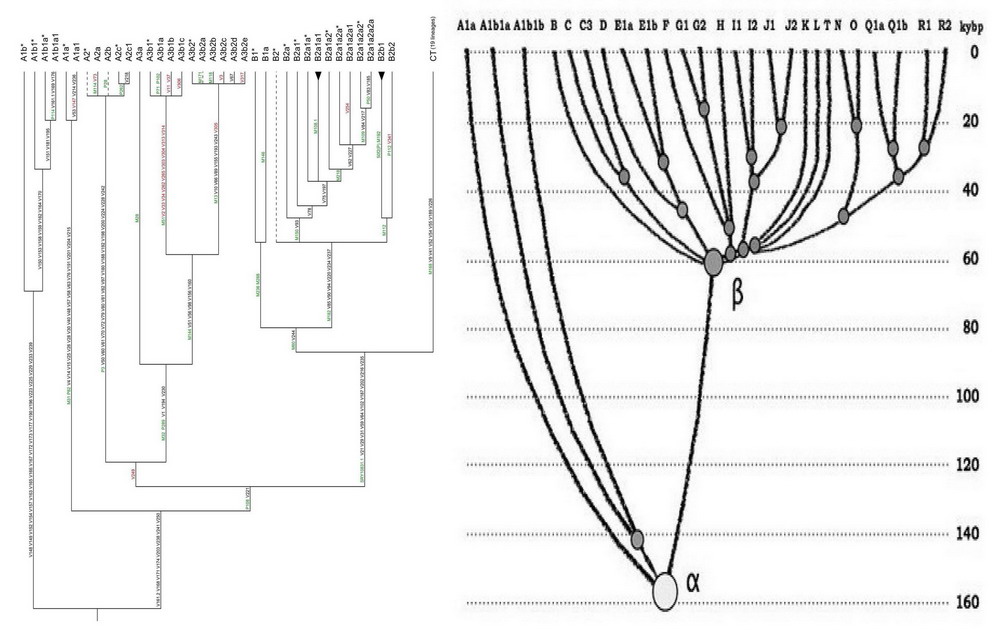 Сопоставление деревьев гаплогрупп и субкладов «африканской» группы (левая часть обоих деревьев) и «неафриканской» (одна линия сводной гаплогруппы СТ на левом дереве и куст гаплогрупп ВТ на правом дереве). Левое дерево (Scozzari et al) опубликовано в ноябре 2012 года, правое дерево (Klyosov, Rozanskii) опубликовано в мае 2012 года. Левое дерево показывает, что серия «африканских» гаплогрупп трижды последовательно отделяется и уходит в сторону от неафриканских, и что неафриканская ДНК-линия СТ (вертикальная линия справа) не происходит от «африканских». Правое дерево показывает те же отщепления «африканских» гаплогрупп от неафриканских (куст гаплогрупп справа), и указывает, что расхождение дерева на африканские и неафриканские гаплогруппа произошли примерно 160 тысяч лет назад. Картинка кликабельна. Оба дерева не показывают недавно обнаруженную гаплогруппу А00, «возраст» которой не менее 200 тысяч лет. Она приведена на следующем рисунке, вместе с изменениями номенклатуры (согласно ISOGG, 2013). Итак, картина в целом прояснилась. Никакого противоречия между деревом Y-хромосомных гаплогрупп человечества, полученным при изучении гаплотипов гаплогрупп от А до Т (Klyosov and Rozhanskii, 2012b, май 2012), и деревом, полученным при геномном изучении Y-хромосомы (Scozzari et al, 2012, ноябрь 2012), нет. Все эти данные, как и другие, показывают глубокое мутационное расхождение между африканскими и неафриканскими ветвями (гаплогруппами, субкладами), и не выявляют «африканское» происхождение анатомически современного человечества. Вместо этого, данные показывают расхождение африканских и неафриканских ДНК-линий примерно 160 тысяч лет назад. Возникает закономерный и недоуменный вопрос – почему, имея все эти данные, авторы исследований продолжают писать, что человечество вышло из Африки относительно недавно, в последние 50-100 тысяч лет? На каком уровне фактического материала или его интерпретаций происходит сбой? Ответить на этот вопрос представляется не менее важным, чем получить ответ, что человечество из Африки не выходило. Посмотрим на упомянутую статью Scozzari et al (2012). В какой момент там появилась фраза об африканском происхождении человека? На чем основана? Эта фраза появляется уже во втором абзаце введения в статью, и сообщает, что гаплогруппа СТ является результатом «недавнего выхода из Африки». В подверждение этого дается ссылка на статью консорциума Проекта «1000 геномов» под названием «Карта вариаций генома человека» (Nature, 2010), в которой про выход из Африки вообще ни слова, как и про гаплогруппу СТ. Понимаете, в чем проблема? Сторонников концепции «выхода из Африки» приходится постоянно ловить за руку, и это продолжается уже более 20 лет. Еще несколько абзацев ниже – опять про «выход из Африки», уже гаплогруппы С, и вообще никакой ссылки. 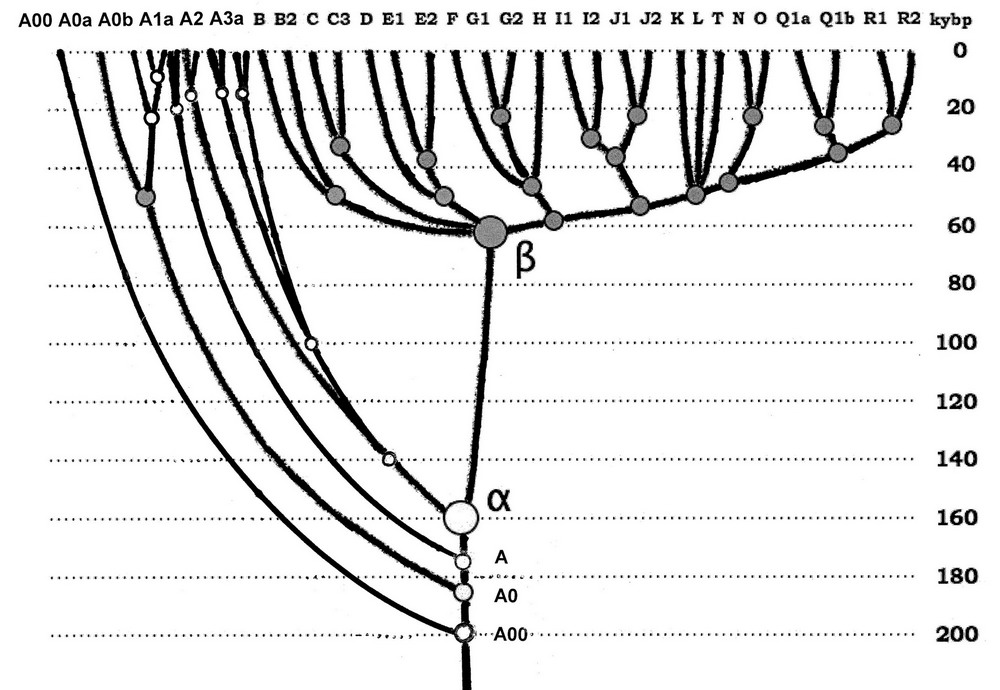 Диаграмма эволюции гаплогрупп современного человечества с добавлением недавно обнаруженной гаплогруппы А00, и с заменой устаревшей номенклатуры 2012 года на номенклатуру 2013 года. На горизонтальной оси – основные гаплогруппы Y-хромосомы человечества, на вертикальной – абсолютная шкала времени. Общий предок альфа-гаплогруппы (гаплогруппы A1b в нынешней классификации) жил примерно 160 тысяч лет назад, общий предок бета-гаплогруппы (или предковой для гаплогрупп от В до Т) – 64±6 тысяч лет назад (из статьи A.A. Klyosov and I.L. Rozhanskii, Advances in Anthropology, 2012b). Идем по статье (Scozzari et al, 2012) дальше. Описывается гаплогруппа A1b (самая верхняя линия на рисунке, взятом из этой статьи, которая первой отошла от остальной части дерева, и по новой номенклатуре называется гаплогруппой A0). Она же отходит влево от дерева на обновлённой диаграмме эволюции гаплогрупп современного человечества примерно 180 тысяч лет назад. Сообщается, что людей с такой мутацией (Р114) найдено очень мало, всего трое из Камеруна, из них один – в данной работе. Мой коментарий – очень хорошо, у меня лично нет больших сомнений, что гаплогруппа А0 и ее ветви – африканские. Но мы от них не произошли, что дерево и показывает. Далее, авторы сообщают, что в Нигере нашли двух человек гаплогруппы А1а – вторая линия в верхней части рисунка из статьи (Scozzari et al, 2012), тоже «африканская». Мой комментарий – и с этим нет никаких проблем. Неафриканских потомков от них тоже нет, согласно тому же рисунку. Далее, гаплогруппа А2, то есть по нынешней номенклатуре А1b1a, третья по счету линия на рисунке из статьи. Авторы сообщают, что носители этой гаплогруппы почти все говорят на щелкающих языках южной Африки, а также являются пигмеями центральной Африки. Авторы нашли троих носителей этой гаплогруппы в Южной Африке. Мой комментарий – замечательно, нет проблем, это вполне африканская линия, и неафриканцы от нее не происходят, как показывает тот же рисунок. В отношении гаплогруппы А3, то есть по нынешней номенклатуре A1b1b-M13, авторы нашли десять носителей этой гаплогруппы – в Эфиопии, Кении и Южной Африке. Еще 28 человек, обладателей этой гаплогруппы из европейских стран и Саудовской Аравии, приведены в Проекте гаплогруппы А, на который выше дан линк. Но пусть эта гаплогруппа считается африканской, все равно неафриканцы от нее не происходят, как видно из рисунка статьи (Scozzari et al, 2012). Гаплогруппу В авторы отправляют к югу от Сахары, а также по всей Африке – центральной, восточной и южной. Мы уже показывали выше, что гаплотипы гаплогруппы В чрезвычайно удалены от «африканских», и явно имеют другое происхождение, родственное неафриканским гаплогруппам. Но это в данном случае не имеет значения, так как «неафриканские» ДНК-линии гаплогруппы СТ не выходят из гаплогруппы В. Они с ней имеют общего предка – гаплогруппу ВТ. И какой же после всего этого делают вывод авторы статьи (Scozzari et al, 2012)? Вы, читатель, будете смеяться, но вывод такой, что гаплогруппы СТ вышли из Африки, и не только из Африки вообще, а из ее северо-западного региона. Именно там, по словам авторов, находятся «истоки разнообразия Y-хромосомы человечества». Как, откуда? Ведь даже беглый взгляд на дерево гаплотипов показывает, что в основе СТ нет африканских источников. Все они – в верхней части диаграммы. А вот так. На этот счет в Америке есть поговорка – «не путайте меня фактами, я уже настроился». Как читатель уже понял, и понял давно, «выход человечества из Африки» стал форменной религией, основанной, как и положено религии, на вере, и аргументы там уже почти бесполезны. Об этом же и другая относительно недавняя статья (Cruciani et al, 2011), в которой уже в названии стоит «происхождение разнообразия в Африке». На каком научном основании? Да все на том же – показали, что африканские Y-хромосомные линии древнее, чем линии неафриканцев. Опять в соответствии с диаграммой выше. Их дерево гаплогрупп почти такое же, как и на рисунке из статьи (Scozzari et al, 2012), но с датировками – 142 тысяч лет назад в сторону от эволюционного Y-хромосомного ствола отошла ветвь А1b (A0 в новой классификации), затем примерно 108 тысяч лет назад отошла ветвь А1а, затем, 105 тысяч лет назад, ветвь А2, затем, те же 105 тысяч лет назад, ветвь А3, которые авторы считают африканскими, поскольку они найдены у четырех африканцев, у которых эти гаплогруппы определяли – и пусть считают, и только затем, 75 тысяч лет назад, отошла ветвь ВТ и далее, 39 тысяч лет назад, ветвь СТ, уже общепризнанно неафриканская. Ни ВТ, ни СТ от «африканских» линий не происходят. Но поскольку точки ветвления авторами названы буквой «А» (А1а-Т, А2-Т), что автоматически принимается за «африканские», то вот и «происхождение из Африки». Поражает такой тип прямолинейного мышления популяционных генетиков. Хорошо, это – мужская, Y-хромосома. Ясно, что соответствующие экспериментальные данные не показывают никакого выхода из Африки. Нет в основе неафриканских гаплогрупп африканских снип-мутаций. Неафриканские гаплотипы исключительно удалены от африканских. Как признают практически все источники, между ними – огромный разрыв, но авторы дальше не идут. Закрыв глаза, повторяют как мантру – «вышли из Африки». Последний раз редактировалось Chugunka; 19.08.2021 в 10:23.
|
|
#5
22.03.2014, 11:42
|
||||
|
||||
|
Перекрестная проверка диаграммы с помощью снипов (SNP)
Существует еще один подход к проверке топологии дерева, представленного на диаграмме чуть выше, с помощью снипов. Дело в том, что в ходе эволюционного развития Y-хромосомы в ней накапливаются практически необратимые мутации, так называемые SNP (Single Nucleotide Polymorphism), или снипы. Чем длиннее переход в диаграмме, тем больше вероятность снипа, тем их больше накапливается в Y-хромосоме (и в остальных хромосомах, но в данном случае мы рассматриваем только Y-хромосому. Самая протяженная эволюционная линия – гаплогруппы А00, она самая архаичная, поэтому в носителях этой гаплогруппы должно обнаружиться максимальное количество снипов. На втором месте по протяженности – гаплогруппа А0, на третьем – линии гаплогруппы А (А1а). Расстояние между альфа- и бета-гаплогруппами (то есть от А1b до ВТ) должно быть относительно небольшим (по сравнению с протяженностью линий А00 и А0), и далее снипы уже накапливаются при переходе от ВТ по линиям конкретных, уже более современных гаплогрупп. Так и оказалось, в подтверждение диаграммы. Я сейчас приведу список снипов по каждой из перечисленных гаплогрупп – с одной стороны, создав некий компактный справочник, чтобы с ним можно было работать, кому нужно, а с другой, чтобы не просто ограничиться на словах неким числом, которое проверить трудно. Правда, эти числа снипов не являются окончательными – время от времени открывают новые снипы. Далее, поскольку снипы появляются неупорядоченно, то мы имеем дело со статистикой, а не с абсолютными, незыблемыми числами. Так что список ниже и число снипов по каждой гаплогруппе находятся в развитии, хотя общее представление дают. Итак – гаплогруппа А00, самая древняя, самая архаичная, ее эволюционная линия на диаграмме самая протяженная. Эти снипы выявили в племени Mbo (Мбо, русскими буквами) в африканском Камеруне: AF4, AF5, AF7, AF8, AF9, AF10, AF13, L990, L1086, L1087, L1088, L1091, L1092, L1094, L1096, L1097, L1100, L1102, L1103, L1104, L1106, L1107, L1108, L1109, L1110, L1111, L1113, L1114, L1115, L1117, L1119, L1122, L1126, L1131, L1133, L1134, L1138, L1139, L1140, L1141, L1144, L1146, L1147, L1148, L1149, L1151, L1152, L1154, L1156, L1157, L1158, L1159, L1160, L1161, L1163, L1233, L1234, L1236, L1284. Всего в гаплогруппе А00 – 59 снипов. Поскольку возраст гаплогруппы А00 оценивается примерно в 210 тысяч лет, то отсюда можно предположить, что снип-мутация происходит в среднем раз в 3600 лет. Далее, как мы уже знаем, на стволе дерева гаплогрупп (диаграмма выше) появилась гаплогруппа А0-Т, от нее отошли гаплогруппы А0 и А1, последняя продолжила ствол дерева. А0 сейчас живут в основном в Африке. Ни одной снип-мутации из списка А00 у А0-Т нет. То есть А0-Т не произошла от африканской гаплогруппы А00. Нет оснований считать гаплогруппу А0-Т африканской. Но она – предок африканской линии А0, и нас, неафриканцев (предки которых прошли через гаплогруппу ВТ, далее СТ, и так далее). Снипы гаплогруппы А0-Т, их 32: AF3, L1085, L1089, L1090, L1093, L1095, L1098, L1099, L1101, L1105, L1116, L1118, L1120, L1121, L1123, L1124, L1125, L1127, L1128, L1129, L1130, L1132, L1135, L1136, L1137, L1142, L1143, L1145, L1150, L1155, L1235, L1273 Гаплогруппа А0 имеет следующие снипы, их 51: L529.2, L896, L982, L984, L990, L991, L993, L995, L996, L997, L998, L999, L1000, L1001, L1006, L1008, L1010, L1011, L1012, L1015, L1016, L1017, L1018, L1055, L1073, L1075, L1076, L1077, L1078, L1080, V148, V149, V152, V154, V157, V163, V164, V165, V166, V167, V172, V173, V176, V177, V190, V196, V223, V225, V229, V233, V239 Как видно, в гаплогруппе А0 на 8 снипов меньше, чем в А00, то есть она примерно на 30 тысяч лет моложе. И действительно, возраст гаплогруппы А0 оценивают в 180 тысяч лет, на 30 тысяч лет меньше, чем возраст гаплогруппы А00. В гаплогруппе А1 пока выявили 21 снип: L985, L986, L989, L1002, L1003, L1004, L1005, L1009, L1013, L1053, L1084, L1112, L1153, P305, V161.2, V168, V171, V174, V238, V241, V250 В отличие от братской гаплогруппы А0, носители А1, видимо, не дожили до настоящего времени. Возможно, они и попали в тот катаклизм, который привел к бутылочному горлышку неафриканской популяции мира. Для оценки суммарного времени жизни гаплогрупп А0-Т, А1 и A1b (в последней – всего две снип-мутации, Р108 и V221) следует сложить число их снип-мутаций, получается 32+21+2 = 55 мутаций, что примерно эквивалентно 198 тысяч лет. Наконец, в гаплогруппе ВТ – 30 снип-мутаций: L413, L418, L438, L440, L604, L957, L962, L969, L970, L971, L977, L1060, L1061, L1062, M42, M91, M94, M139, M299, P97, SRY10831.1, V21, V29, V59, V64, V102, V187, V202, V216, V235 Это дает 108 тысяч лет эволюции бета-гаплогруппы от времени расхождения с африканской гаплогруппой A1b1 (160±12 тысяч лет назад) до прохождения бутылочного горлышка популяции 64±6 тысяч лет назад. Это и есть пропавшие 108 тысяч лет на диаграмме (дистанция между альфа- и бета-гаплогруппой). Важно то, что эти пропавшие тысячелетия в эволюции неафриканских ДНК-генеалогических линий воспроизводятся как при анализе гаплотипов (на основании чего и была построена диаграмма выше), так и при анализе снип-мутаций. Вот это и есть перекрестная проверка диаграммы. Почему мы не видим этих пропавших людей в Евразии – неизвестно. С другой стороны, известны скелетные остатки Homo sapiens с датировкой между 160 и 60 тысяч лет, обнаруженные на Ближнем Востоке, но их гаплогруппа не проверялась. Если окажется гаплогруппа BT c какими-то из мутаций, приведенных в списке выше – загадка будет окончательно решена. Неафриканские мтДНК не происходят из африканских А что на этот счет говорят данные по женской, мтДНК? Ну, говорят сторонники «выхода из Африки», там-то тем более ясно. Неафриканские мтДНК «все происходят из африканских». Так ли это? Взглянем на недавнюю статью известного генетика Д. Бехара (Behar et al, 2012), в которой была проведена принципиальная ревизия характера представления последовательности мтДНК. На последующем рисунке видно, что в самом начале эволюционного дерева мтДНК «современного человека» происходит расхождение гаплогрупп на L0 (ветвь справа) и гаплогруппы L1-L6 (ветвь слева), из которых далее происходят все последующие гаплогруппы. Гаплогруппа L0 на самом деле представляет большую серию из полусотни древних африканских гаплогрупп, в основном найденных к югу от Сахары – среди койсанского населения Южной Африки, а также в Эфиопии и Танзании (Восточная Африка), в Мозамбике (Юго-Восточная Африка), и среди пигмеев. Все остальные мтДНК происходят, как принято большинством специалистов, из гаплогруппы L3, возраст которой по оценкам примерно 60-70 тысяч лет, то есть такой же, как гаплогруппы ВТ Y-хромосомы. То есть вполне вероятно, что гаплогруппа L3 не вышла из Африки, а, напротив, пришла в Африку вместе с носителями Y-хромосомы, например, гаплогруппы ВТ. Какой же вывод делает Бехар с соавторами? Естественно, «человечество вышло из Африки». Глубокое расхождение африканской гаплогруппы L0 с остальными в их статье даже не обсуждается, хотя из данных статьи сразу видно, что африканская гаплогруппа L0 не является предковой по отношению ко всем остальным гаплогруппам мтДНК. 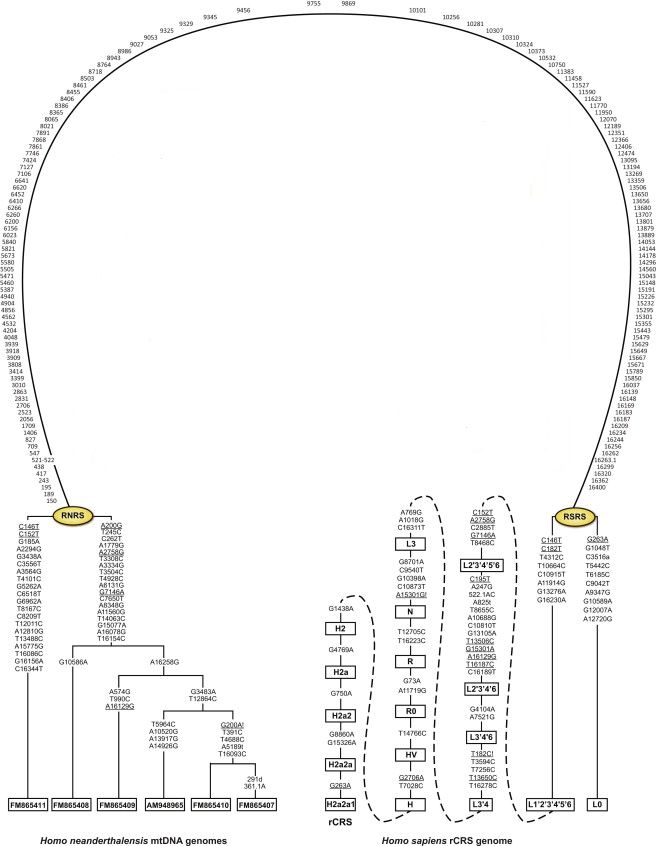 Схематическое представление мтДНК человека, показывающее мутационную связь между мтДНК неандертальца (слева) и мтДНК человека разумного (справа). В овале слева и справа сокращения RNRS и RSRS означают соответственно «реконструированная референсная последовательность неандертальца» и «реконструированная референсная последовательность человека». Из работы (Behar et al.,2012). Обратите внимание на резкое расхождение в начале референсной последовательности человека (справа) цепи мутаций на африканскую гаплогруппу L0 (справа внизу), и цепочку всех остальных гаплогрупп мтДНК. Картинка кликабельна. На самом деле, расхождение африканской гаплогруппы L0 (возраст 150-170 тысяч лет) и изначально неафриканских гаплогрупп L1-L6, с последующим приходом гаплогруппы L3 (возраст 60-70 тысяч лет) в Африку оказывается практически совпадающим с расхождением Y-хромосомных гаплогрупп африканской серии А (возраст 160 тысяч лет) и изначально неафриканских ВТ (возраст 64 тысячи лет) с последующим приходом гаплогруппы В в Африку. Так что и в отношении мтДНК концепция «выхода из Африки» оказывается совершенно необязательной, и фактически построена на песке. Описание мтДНК в академических статьях и справочниках пестрит словами «возможно», «вероятно», «предполагается», что фактически означает, что данных нет, и что все построено на предположениях. Одна проблема – все эти предположения всегда трактуются только в одну сторону – «выхода из Африки». Вместе с тем накапливается все больше данных, что в древнем прошлом миграции популяций в Африку происходили многократно. Только что вышла статья в журнале Nature (Hayden, 2013), в которой сообщается о двух миграциях из Евразии в койсанские племена в Южной Африке, одна 3 тысячи лет назад в восточную Африку, другая, ее продолжение – 900-1800 лет назад в Южную Африку. Какие гаплогруппы принесли мигранты – не сообщается. Но нет сомнения, что они резко увеличили «генетическое разнообразие» койсан, которое считается максимальным в Африке. Вторая важная особенность этого сообщения – оно указывает на возможность миграций «в Африку», в чем, впрочем, никакого сомнения и не могло быть. Неясно, почему с таким упорством сторонники «выхода человечества из Африки» держатся именно за их односторонний вариант. Впрочем, упорство уменьшается, и вот уже Сара Тишкоф из Пенсильванского университета, одна из наиболее активных защитников «из Африки», уже приветствует новые данные «в Африку», и называет их «имеющими смысл», поскольку «об этом говорят и археологические, и лингвистические исследования» (Nature, 29 августа 2013, стр. 514). Миграции вируса герпеса ошибочно объявлены как якобы подтверждающие «выход человечества из Африки» История с недавней нашумевшей публикацией «Использование филогенетики генома простого вируса герпеса для прослеживания древних миграций человека» является показательной – как исходная установка на якобы выход современного человечества из Африки искажает выводы научной публикации. Напомню, что в статье сопоставляли 31 геномные последовательности простого вируса герпеса в популяциях из Восточной Африки (Кения), Восточной Азии (Китай, Южная Корея, Япония), Северной Америки (США) и Европы (Соединенное Королевство), и обнаружили, что построенное филогенетическое дерево расходится на шесть кластеров. Эти кластеры соответствовали следующим популяциям: I – из 10 образцов 7 были из Сиэттла, один «из США», один из Сан-Франциско, и самый нижний – (номер 17) из Шотландии. II – из 6 образцов один был из Китая, один из США (Хьюстон), два из Южной Кореи и два из Японии. III – два образца из Кении. IV – три образца из Кении. V – семь образцов из Кении. VI – два образца из Кении. Авторы статьи данные «округлили» и сообщили, что первый кластер «объединяет Северную Америку и Европу», второй – Восточную Азию, остальные – «Восточную Африку». Происхождение американских образцов вируса в статье не дано, то есть неясно, получены они от американских индейцев или, например, потомков англосаксов, но поскольку авторы предположили, что выпадающий из даже округленного варианта образец из Хьюстона, который попал к восточно-азиатским вирусам, мог принадлежать американскому индейцу, то становится ясно, что остальные северо-американские образцы – скорее всего от потомков европейцев. Тем не менее, авторы заключили, что эти кластеры отражают «глобальные миграции (древнего) человека», и подтверждают выход современного человечества из Африки (supports… the «out of Africa» theory of human evolution). Основная причина – как обычно, аргумент «разнообразия», а именно что «вирусы восточно-африканского происхождения имеют наибольшее разнообразие и образуют четыре из шести кластеров». Давайте, как мы уже делали выше, посмотрим, откуда появляется это «разнообразие». А появляется оно опять от расхождения популяций в разные стороны – одни уходят в Африку, другие не уходят. Но они опять не происходят друг от друга, они каждый раз происходят от более древнего общего предка, который в Африке вполне мог не жить. То есть опять повторяются ситуации, описанные ранее в этой статье. Никакого «африканского происхождения» показанные данные не дают.  Филогенетическое дерево, показывающее динамику штаммов простого вируса герпеса HSV-1 и его каскадное расхождение на шесть ветвей, как насчитывают в работе (Kolb et al., 2013), которые, по мнению тех же авторов, следуют определенной географии: ветвь I – Европа/Северная Америка (штамм 17 – из Глазго, остальные – Сиэтл, Сан Франциско, «США»), ветвь 2 – Восточная Азия (Китай, Южная Корея, Япония, и Хьюстон [CША]), ветви III, IV, V и VI – восточно-африканские (все из Кении). Посмотрим на приведенную диаграмму. Слева – первое расхождение от древнего общего предка. В сторону отходят вирусы, которые сегодня находят у кенийцев (два самых верхних образца, кластер VI). От них никакие неафриканские вирусы не происходят. Далее – следующее расхождение популяций вирусов (и, видимо, их носителей-людей). В сторону отходит кластер V, ныне кенийцы. От них никакие неафриканские вирусы не происходят. Далее – опять вилка, очередное расхождение популяций. В сторону отходит опять кенийский вирус, с индексом Е07 на схеме выше. От него неафриканские вирусы опять не происходят. Его объединение с остальными кенийскими образцами в кластер IV – ошибка авторов, нет там общего кластера. Следом – еще две вилки-расхождения популяций, и каждый раз в сторону отходит кенийская группа (кластеры IV и III), от которой европейцы и азиаты не происходят. При последнем расхождении в сторону отходит исключительно неафриканская группа. Она опять не происходит от африканцев. Последнее расхождение – на европейские и восточно-азиатские (за некоторым исключением) линии вирусов герпеса, кластеры I и II. Америка там, судя по всему, не при чем, это не древние обитатели Америки, а относительно недавние переселенцы из Европы. Поразительно, что авторы не видят такого простого объяснения динамики вирусов. Они полностью поглощены тем, как подогнать данные к «выходу из Африки». Зачем, какая цель – понять невозможно. В своем комментарии к статье, ведущий автор Куртис Брандт, профессор медицинской микробиологии и офтальмологии, сообщил, что результаты оказались «ошеломляющими», и далее – «мы нашли, что все африканские изоляты образуют один кластер, все вирусы с Дальнего Востока, Кореи, Японии, Китая вместе образуют другой кластер, и все вирусы из Европы и Америки, за одним исключением, образуют еще один кластер». В общем, невзирая на очевидные натяжки, это верно. Но показывает ли это «африканское происхождение современного человека»? Ясно, что нет, как это объяснено выше. В другом комментарии д-р Брандт поделился вполне откровенно: «мы нашли точно то, что нам сказали молекулярные генетики, которые изучают геном человека, а именно откуда люди произошли (из Африки – АК) и как они расходились по планете». Вот это и есть основная проблема этого и подобных исследований. Им сказали (неверно), они приняли как руководство к действию и нашли именно это. Статья предоставляет еще одну иллюстрацию того, как данные подгоняются под желаемый «выход из Африки». Это то, как авторы рассчитывали времена этого «выхода», базируясь на тех самых предполагаемых 50 тыс. лет назад, и на самом деле никогда не показанных. Это – «консенсусные данные». На самом деле, разные авторы приводят датировки от 27 до 200 тысяч лет назад, как правило, опять же без расчетов, но цифры 50 или 70 тыс. лет назад почему-то выглядят им привлекательными. Недавно, правда, «консенсус» начал смещаться в 100-140 тыс. лет назад, но 50 или 70 тыс. лет по инерции продолжают приводиться. Вот и авторы обсуждаемой статьи их, эти умозрительные датировки, использовали как базовые. Давайте посмотрим, что из этого авторы получили. Это весьма поучительно. Как сообщили авторы статьи, в литературе имеются три существенно различающиеся величины констант скоростей мутаций вируса простого герпеса и других вирусов герпеса, равные 3х10-9, 18.2х10-9, и 30х10-9 мутаций на нуклеотид в год. Применение их для расчетов времен расхождения вируса по популяциям человека, описанным выше, дало бы расхождения по временам в 30 раз. Понятно, что такая неопределенность авторов не устраивала, и они решили фактически подогнать времена исходного расхождения популяций под «консенсусное» время «выхода из Африки», и на первом этапе расчетов подогнать под предполагаемое время расхождения европейской и азиатской популяции, которое взяли как 23-45 тысяч лет назад, процитировав четыре литературных источника на этот счет. Усреднив эти величины, авторы взяли как «референсную» величину 34,000±10,500 лет назад. Правда, вместо «европейской» авторы постоянно упоминали «европейская/северо-американская», хотя ясно, что по сути это европейская – один образец вируса из Шотландии, все другие (в основном из Сиэттла) – скорее всего, потомки переселенцев из Европы. В любом случае эти цифры заниженные, так как разделение гаплотипов Европы и Азии произошло не позднее 55-60 тысяч лет назад, древнейшие костные остатки «анатомически современного человека» в Европе датируются 45 тыс. лет назазд, аборигены пришли в Австралию не позднее 50 тыс. лет назад, но в статье все настолько приблизительное – и датировки, и скорости мутации вируса, что к этому никак нельзя относиться серьезно. Поскольку времена у авторов намного заниженные, то скорости мутации должны были получиться намного завышенные. Так и получилось – подогнанная таким образом константа скорости мутации вируса оказалась намного быстрее известных (точнее, литературных) величин, а именно 134х10-9 мутаций на нуклеотид в год, с верхним и нижним пределами 214х10-9 и 74.8х10-9, соответственно. Используя эту подогнанную величину константы скорости мутации, авторы получили, что исходное расхождение вируса произошло 50.3±16.7 тысяч лет назад, и постановили, что это соответствует «выходу человечества из Африки». Слегка подправленная датировка расхождения европейцев и азиатов по расчетам авторов оказалась 32.8±10.9 тыс. лет назад, а время расхождения единственного китайского образца и единственного образца из Техаса оказалось 15.76±5.3 тысяч лет назад, что авторы отнесли к заселению Америки, «которое состоялось в тот период». Здесь комментарии просто излишни. Все это легло в основу провозглашения авторами, что «впервые показано, что филогенетические данные по вирусу герпеса подтверждают выход человечества из Африки». Как на самом деле показано выше, это не имеет никакого отношения к «выходу человечества из Африки». Более того, оценка величины константы скорости мутации вируса герпеса, проведенная авторами на основании более чем грубых прикидок превышает три другие литературные величины в 4.5 – 45 раз. Никаких перекрестных проверочных тестов по верификации полученной константы скорости мутации авторами не делалось. То, что авторы получили, исходя из их константы, что вирусы герпеса HSV-1 и HSV-2 разошлись 2.184±0.753 миллионов лет назад (обратите внимание на приведенную «точность» в три знака после запятой!), ни о чем не говорит, там могло получиться и 20 миллионов лет назад с тем же успехом, и тоже ничего бы не сказало – это могло быть у макаки, например. Если все-таки более верны литературные данные, то исходное расхождение вируса герпеса могло произойти не 50.3 тысяч лет назад, а 220 тыс. лет – 2.2 миллиона лет назад, и тоже могло быть неплохо интерпретировано в рамках эволюции человека. Так что исходные, экспериментальные данные авторов представляют несомненную ценность, а манипуляции, выводы и интерпретации – никакой ценности не представляют. Так, к сожалению, сейчас часто «делается наука», особенно в области популяционной генетики. Антропологические данные и датировки В последние годы сложилась занятная ситуация, когда многие антропологи выражают сомнение в «африканском происхождении человека», но кивают на генетиков, что мы, мол, ответа не имеем, антропологические данные противоречивы, но вот генетики утверждают, что точно знают, что из Африки, поэтому как мы можем спорить? А те генетики, которые понимают, что генетические данные построены на песке, точнее, на слишком вольных (фантазийных) интерпретациях, кивают на антропологов, что, мол, мы понимаем, что генетические данные слабы и часто просто неверны, но вот антропологи утверждают, что из Африки, и датировки их на это же указывают, поэтому как мы можем спорить? Значит, и у нас все верно. Посмотрим на утверждения, что анатомически современный человек (АСЧ) якобы точно произошел в Африке, и было это около то ли 40-50 тысяч лет назад, то ли 100, то ли 150, то ли 200 тысяч лет назад. Напомним, что АСЧ – это тот, кто не имеет заметных архаичных антропологических особенностей. Сначала набросаем краткое описание обстановки, потом продемонстрируем это на конкретном материале. Если действительно кратко, то имеем пять основных положений: (1) все африканские находки древних костей до примерно 36 тысяч лет назад показывают заметные архаичные особенности; (2) зачастую древние кости настолько фрагментарны, что по ним даже минимальную антропологическую картину воссоздать просто нельзя; (3) часто антропологическая картина костей в Африке и за пределами Африки очень схожа, и потому нельзя сказать, это был выход из Африки или вход в Африку; (4) часто костных остатков вообще нет, и утверждения об «анатомически современных людях» делаются на основании стоянок и каменных орудий, хотя их вполне могли оставить архантропы, то есть архаичные люди, не относящиеся к виду «человек разумный»; (5) датировки древних костей часто настолько под вопросом, что мало кто их воспринимает в буквальном смысле или даже просто серьезно. Начнем с последнего. К сожалению, радиоуглеродные датировки на временах больше примерно 40 тысяч лет назад уже почти не работают, и недавний рекорд по датировке – 60 тысяч лет назад. Причина проста – период полураспада радиоактивного изотопа 14С составляет 5730 лет, то есть 40 тысяч лет – это семь периодов полураспада, а 60 тысяч лет – это более десяти периодов полураспада. Метод основан на измерении соотношения в биологических образцах содержания стабильного изотопа 12С (и немного 13С, почти в сто раз меньше по сравнению с содержанием 12С) и радиоактивного 14С (при его исходном содержании в количестве одной десятимиллиардной процента), которое со временем падает, с тем самым периодом полураспада. За 60 тысяч лет его содержание уменьшается от исходных 10-10% в 210 раза, то есть еще в 1024 раза. Современные приборы такие уровни радиации уже не ловят, во всяком случае приборы, которые находятся в пользовании археологов. Это – примерно 1 щелчок в час на грамм тестируемого углерода. Обычный фон значительно выше. Это открывает «широкие возможности» для ошибок, и не только ошибок. Вспомним нашумевшую (в узких кругах) историю с разжалованием и увольнением со скандалом немецкого антрополога Рейнера Протча (Protsch) за систематические, как оказалось, фальсификации датировок древних (и вовсе не древних) костей. После проверки датировка Протча в 36000 лет назад оказалось 7500 лет назад, датировка 21300 лет назад оказалась 2300 лет назад, а скелет, датированный им 29400 лет назад, оказался останками человека, который умер в 1750 году, за 255 лет до измерений (газета Гардиан). Об этом же писал и известный международный журнал «Археология». Подобных историй немало в археологии, но если и отбросить явных фальсификаторов, которых, конечно, только единицы, то вероятность ошибок в любом случае велика. Особенно когда удревнить очень хочется, и тем самым войти в историю археологии, а то и культуры человечества в целом. Надо сказать, что особенно древние биологические находки датируются уже другими методами, как, например, аргоновым, по отношению содержания 40Ar/39Ar. В целом же, нет никаких антропологических или археологических доказательств «происхождения современного человека в Африке», как и нет никаких доказательств, что каменные «инструменты» и «индустрии», найденные в Европе или в Евразии в целом, были привнесены с территории к югу от Сахары. Все заявления об обнаружениях костных останков «анатомически современного человека» давностью больше 50 тысяч лет, и тем более больше 150 тысяч лет, и тем более к югу от Сахары, являются просто искаженными или неверными с самого начала. Довольно полный обзор на эту тему будет вскоре опубликован известным австралийским антропологом Робертом Беднариком (в печати, Advances in Anthropology). Об отсутствии таких находок к югу от Сахары свидетельствуют ряд работ, например (Grine et al, 2007; Grine et al, 2010). Все известные находки костей такой древности носят явные архаичные особенности, начиная с Omo Kibish 1 (195 тысяч лет назад, Эфиопия, найдены фрагменты черепных костей, лицевых костей мало), Omo-2 (показан ряд примитивных, архаичных особенностей), Herto (154-160 тыс лет назад, очень архаичное костное строение, сильно отличающееся от АСЧ), в общем, все скелетные останки, датируемые 100-200 тысяч лет назад, и фактически ранее 35 тысяч лет назад, являются архаичными (Rightmire, 2009). Многие вообще не имеют сохранившихся лицевых костей. Даже череп Hofmeyr из Южной Африки, датируемый 36 тысяч лет назад, имеет архаичные особенности (Grine et al, 2007; Rightmire, 2009; Tattersall, 2009). Американский антрополог Райтмайр сообщил – «ни ископаемые Herto, ни другие из позднего Плейстоцена, как Klasies River в Южной Африке или Skhul/Qafzeh в Израиле, не имеют аналогий с современными популяциями. Их черепа являются робастными, и только начиная с ∼ 35000 лет назад начинают появляться люди современной анатомической морфологии» (Rightmire, 2009). Он полагает, что «анатомически современные люди» эволюционировали в Африке, хотя этот процесс «плохо понят». Ему вторит Майкл Хаммер (Hammer et al, 2011) – «ископаемые гоминины, показывающие комбинацию архаичных и более современных особенностей, постоянно встречаются в Африке к югу от Сахары и на Ближнем Востоке вплоть до примерно 35 тысяч лет назад». Таким образом, постоянные ссылки сторонников концепции «из Африки», что в Африке обнаружены костные остатки анатомически современных людей с датировками между 160 и 200 тысяч лет назад, являются ложными. Манипуляции и искажения продолжаются и здесь. Майкл Хаммер, в недавнем прошлом активный защитник концепции «из Африки», стал выражать сомнения в выходе «из Африки», но стал это выражать уже в 2013 году, когда стало ясно, что ситуация с «выходом из Африки» запуталась донельзя. Хаммер заканчивает свою статью в журнале «Сайнтифик Америкэн» (май 2013 года) следующими словами: «Много узлов остались нераспутанными. Но одно ясно – корни современного человека идут не только к единственной предковой популяции в Африке, но к популяциям Старого Света» (то есть Европы или Евразии – АК). И это представляется вполне разумным. Изучая историю древнего мира, мы постоянно встречается с мобильными миграциями, в том числе и весьма дальними миграциями. Если по версии сторонников «выхода из Африки» человек всего за 10 тысяч лет достиг Австралии, то трудно себе представить, что он сиднем сидел 200 тысяч лет и не разошелся по всему миру, в том числе и в Африку и из Африки, причем многократно. Как получилось, что концепция одностороннего «выхода из Африки» так агрессивно навязывалась обществу и так быстро им овладела, должно вызывать тревогу – и в отношении агрессивного, безапелляционного навязывания, и в отношении столь безвольного восприятия общественностью. И ведь что особенно должно тревожить – ведь никаких надежных оснований у «выхода из Африки» не было и нет. Резюмируя, повторим еще раз – наши предки из Африки в последние 200 тысяч лет не выходили. Иначе говоря, те, кто из Африки выходили, не породили современное неафриканское человечество. Это показывает весь комплекс полученные данных – и генетика, и антропология, и археология, и ДНК-генеалогия. На самом деле и не только в последние 200 тысяч лет, но и раньше. Изучение ДНК ископаемых костей нендертальцев показало наличие в них рецептора меланокортина (MCR1), причем в варианте, который задает светлую кожу и рыжие волосы (Lalueza-Fox et al, 2007). Авторы полагают, что неандертальцы по цвету волос примерно соответствовали современным европейцам – от темноволосых до блондинов. К тому же не обнаружено никаких показателей, что неандертальцы были негроидами. Действительно, в Африке следов неандертальцев не выявлено. А поскольку наши ближайшие предки с неандертальцами были общие, поскольку неандерталец – наш племянник, то и «отец» неандертальца и «брат» нашего «отца» тоже, скорее всего, имел светлую кожу и жил не в Африке. Было это где-то в интервале 300-600 тысяч лет назад. Остается, впрочем, неразгаданным, как наши светлокожие братья, с которыми мы разошлись примерно 160 тысяч лет назад, выжили, попав в Африку, и как приобрели темный цвет кожи, но ответ найдется тоже в области генетики, в регуляции биосинтеза меланина. Но это уже другая история. Литература Анатолий А. Клёсов, профессор, доктор химических наук
|
 |
| Здесь присутствуют: 1 (пользователей: 0 , гостей: 1) | |
|
|





 Линейный вид
Линейный вид

